Refine listing
Actions for selected content:
32 results in 37Gxx
The cyclicity of the period annulus of a reversible quadratic system
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 152 / Issue 2 / April 2022
- Published online by Cambridge University Press:
- 10 February 2021, pp. 281-290
- Print publication:
- April 2022
-
- Article
- Export citation
-
We prove that perturbing the periodic annulus of the reversible quadratic polynomial differential system
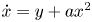 $\dot x=y+ax^2$,
$\dot x=y+ax^2$, 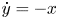 $\dot y=-x$ with a ≠ 0 inside the class of all quadratic polynomial differential systems we can obtain at most two limit cycles, including their multiplicities. Since the first integral of the unperturbed system contains an exponential function, the traditional methods cannot be applied, except in Figuerasa, Tucker and Villadelprat (2013, J. Diff. Equ., 254, 3647–3663) a computer-assisted method was used. In this paper, we provide a method for studying the problem. This is also the first purely mathematical proof of the conjecture formulated by Dumortier and Roussarie (2009, Discrete Contin. Dyn. Syst., 2, 723–781) for q ⩽ 2. The method may be used in other problems.
$\dot y=-x$ with a ≠ 0 inside the class of all quadratic polynomial differential systems we can obtain at most two limit cycles, including their multiplicities. Since the first integral of the unperturbed system contains an exponential function, the traditional methods cannot be applied, except in Figuerasa, Tucker and Villadelprat (2013, J. Diff. Equ., 254, 3647–3663) a computer-assisted method was used. In this paper, we provide a method for studying the problem. This is also the first purely mathematical proof of the conjecture formulated by Dumortier and Roussarie (2009, Discrete Contin. Dyn. Syst., 2, 723–781) for q ⩽ 2. The method may be used in other problems.
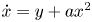
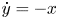
On the birth and death of algebraic limit cycles in quadratic differential systems
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 32 / Issue 2 / April 2021
- Published online by Cambridge University Press:
- 26 May 2020, pp. 317-336
-
- Article
- Export citation
Phase portraits of the quadratic polynomial Liénard differential systems
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 151 / Issue 1 / February 2021
- Published online by Cambridge University Press:
- 04 March 2020, pp. 202-216
- Print publication:
- February 2021
-
- Article
- Export citation
-
We classify the global phase portraits in the Poincaré disc of the quadratic polynomial Liénard differential systems
where (x, y) ∈ ℝ2 are the variables and a,b,c,d,e are real parameters.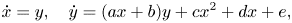 \dot{x}=y, \quad \dot{y}=(ax+b)y+cx^2+dx+e,
\dot{x}=y, \quad \dot{y}=(ax+b)y+cx^2+dx+e,
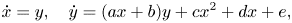
Two components is too simple: an example of oscillatory Fisher–KPP system with three components
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 150 / Issue 6 / December 2020
- Published online by Cambridge University Press:
- 24 September 2019, pp. 3097-3120
- Print publication:
- December 2020
-
- Article
- Export citation
Parametric rigidity of real families of conformal diffeomorphisms tangent to x→−x
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 149 / Issue 1 / February 2019
- Published online by Cambridge University Press:
- 30 August 2018, pp. 261-277
- Print publication:
- February 2019
-
- Article
- Export citation
Integrable zero-Hopf singularities and three-dimensional centres
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 148 / Issue 2 / April 2018
- Published online by Cambridge University Press:
- 17 October 2017, pp. 327-340
- Print publication:
- April 2018
-
- Article
- Export citation
SINGULAR COTANGENT MODEL
- Part of
-
- Journal:
- Glasgow Mathematical Journal / Volume 57 / Issue 2 / May 2015
- Published online by Cambridge University Press:
- 18 December 2014, pp. 415-430
- Print publication:
- May 2015
-
- Article
-
- You have access
- Export citation
HOPF BIFURCATION ANALYSIS FOR A RATIO-DEPENDENT PREDATOR–PREY SYSTEM INVOLVING TWO DELAYS
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 55 / Issue 3 / January 2014
- Published online by Cambridge University Press:
- 05 June 2014, pp. 214-231
-
- Article
-
- You have access
- Export citation
DYNAMICAL SYSTEMS ANALYSIS OF A MODEL DESCRIBING TASMANIAN DEVIL FACIAL TUMOUR DISEASE
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 54 / Issue 1-2 / October 2012
- Published online by Cambridge University Press:
- 18 March 2013, pp. 89-107
-
- Article
-
- You have access
- Export citation
ON THE NUMBER OF LIMIT CYCLES IN PERTURBATIONS OF A QUADRATIC REVERSIBLE CENTER
- Part of
-
- Journal:
- Journal of the Australian Mathematical Society / Volume 92 / Issue 3 / June 2012
- Published online by Cambridge University Press:
- 11 October 2012, pp. 409-423
- Print publication:
- June 2012
-
- Article
-
- You have access
- Export citation
The SIS Great Circle Epidemic Model
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 45 / Issue 2 / June 2008
- Published online by Cambridge University Press:
- 14 July 2016, pp. 513-530
- Print publication:
- June 2008
-
- Article
-
- You have access
- Export citation
Stochastic and deterministic analysis of SIS household epidemics
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 38 / Issue 4 / December 2006
- Published online by Cambridge University Press:
- 08 September 2016, pp. 943-968
- Print publication:
- December 2006
-
- Article
-
- You have access
- Export citation
