Refine listing
Actions for selected content:
82 results in 34Kxx
Periodic dynamics of a general switching dynamical system
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 08 August 2025, pp. 1-15
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Optimal stopping zero-sum games in continuous hidden Markov models
- Part of
-
- Journal:
- Advances in Applied Probability , First View
- Published online by Cambridge University Press:
- 28 July 2025, pp. 1-44
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
A DELAY DYNAMICAL SYSTEM’S PERSPECTIVE ON THE GLUCOSE–INSULIN REGULATORY RESPONSE TO ON–OFF GLUCOSE INFUSION
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 67 / 2025
- Published online by Cambridge University Press:
- 22 July 2025, e26
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Impact of prey-taxis on a harvested intraguild predation predator–prey model
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 10 March 2025, pp. 1-38
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Two dynamical approaches to the notion of exponential separation for random systems of delay differential equations
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 24 February 2025, pp. 1-39
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
This paper deals with the exponential separation of type II, an important concept for random systems of differential equations with delay, introduced in Mierczyński et al. [18]. Two different approaches to its existence are presented. The state space X will be a separable ordered Banach space with
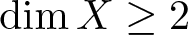 $\dim X\geq 2$, dual space
$\dim X\geq 2$, dual space 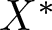 $X^{*}$, and positive cone
$X^{*}$, and positive cone 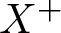 $X^+$ normal and reproducing. In both cases, appropriate cooperativity and irreducibility conditions are assumed to provide a family of generalized Floquet subspaces. If in addition
$X^+$ normal and reproducing. In both cases, appropriate cooperativity and irreducibility conditions are assumed to provide a family of generalized Floquet subspaces. If in addition 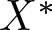 $X^*$ is also separable, one obtains an exponential separation of type II. When this is not the case, but there is an Oseledets decomposition for the continuous semiflow, the same result holds. Detailed examples are given for all the situations, including also a case where the cone is not normal.
$X^*$ is also separable, one obtains an exponential separation of type II. When this is not the case, but there is an Oseledets decomposition for the continuous semiflow, the same result holds. Detailed examples are given for all the situations, including also a case where the cone is not normal.
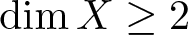
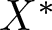
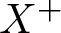
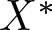
Hopf bifurcations for a delayed discrete single population patch model in advective environments
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 11 November 2024, pp. 705-737
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
A diffusive plant-sulphide model: spatio-temporal dynamics contrast between discrete and distributed delay
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 28 October 2024, pp. 811-828
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
The dynamical analysis of a nonlocal predator–prey model with cannibalism
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 35 / Issue 5 / October 2024
- Published online by Cambridge University Press:
- 29 January 2024, pp. 707-731
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Branching patterns of wave trains in mass-in-mass lattices
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 155 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 11 January 2024, pp. 1240-1266
- Print publication:
- August 2025
-
- Article
- Export citation
-
We investigate the existence and branching patterns of wave trains in the mass-in-mass (MiM) lattice, which is a variant of the Fermi–Pasta–Ulam (FPU) lattice. In contrast to FPU lattice, we have to solve coupled advance-delay differential equations, which are reduced to a finite-dimensional bifurcation equation with an inherited Hamiltonian structure by applying a Lyapunov–Schmidt reduction and invariant theory. We establish a link between the MiM lattice and the monatomic FPU lattice. That is, the monochromatic and bichromatic wave trains persist near $\mu =0$
 in the nonresonance case and in the resonance case $p:q$
in the nonresonance case and in the resonance case $p:q$ where $q$
where $q$ is not an integer multiple of $p$
is not an integer multiple of $p$ . Furthermore, we obtain the multiplicity of bichromatic wave trains in $p:q$
. Furthermore, we obtain the multiplicity of bichromatic wave trains in $p:q$ resonance where $q$
resonance where $q$ is an integer multiple of $p$
is an integer multiple of $p$ , based on the singular theorem.
, based on the singular theorem.







Long-time dynamics and semi-wave of a delayed nonlocal epidemic model with free boundaries
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 155 / Issue 2 / April 2025
- Published online by Cambridge University Press:
- 05 October 2023, pp. 564-610
- Print publication:
- April 2025
-
- Article
- Export citation
Periodic solutions of four-order degenerate differential equations with finite delay in vector-valued function spaces
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 155 / Issue 2 / April 2025
- Published online by Cambridge University Press:
- 14 September 2023, pp. 395-412
- Print publication:
- April 2025
-
- Article
- Export citation
-
In this paper, we mainly investigate the well-posedness of the four-order degenerate differential equation ($P_4$
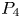 ): $(Mu)''''(t) + \alpha (Lu)'''(t) + (Lu)''(t)$
): $(Mu)''''(t) + \alpha (Lu)'''(t) + (Lu)''(t)$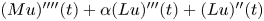 $=\beta Au(t) + \gamma Bu'(t) + Gu'_t + Fu_t + f(t),\,( t\in [0,\,2\pi ])$
$=\beta Au(t) + \gamma Bu'(t) + Gu'_t + Fu_t + f(t),\,( t\in [0,\,2\pi ])$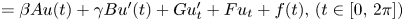 in periodic Lebesgue–Bochner spaces $L^p(\mathbb {T}; X)$
in periodic Lebesgue–Bochner spaces $L^p(\mathbb {T}; X)$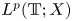 and periodic Besov spaces $B_{p,q}^s\;(\mathbb {T}; X)$
and periodic Besov spaces $B_{p,q}^s\;(\mathbb {T}; X)$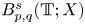 , where $A$
, where $A$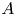 , $B$
, $B$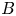 , $L$
, $L$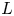 and $M$
and $M$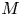 are closed linear operators on a Banach space $X$
are closed linear operators on a Banach space $X$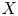 such that $D(A)\cap D(B)\subset D(M)\cap D(L)$
such that $D(A)\cap D(B)\subset D(M)\cap D(L)$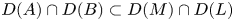 and $\alpha,\,\beta,\,\gamma \in \mathbb {C}$
and $\alpha,\,\beta,\,\gamma \in \mathbb {C}$ , $G$
, $G$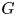 and $F$
and $F$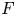 are bounded linear operators from $L^p([-2\pi,\,0];X)$
are bounded linear operators from $L^p([-2\pi,\,0];X)$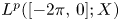 (respectively $B_{p,q}^s([-2\pi,\,0];X)$
(respectively $B_{p,q}^s([-2\pi,\,0];X)$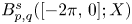 ) into $X$
) into $X$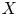 , $u_t(\cdot ) = u(t+\cdot )$
, $u_t(\cdot ) = u(t+\cdot )$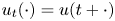 and $u'_t(\cdot ) = u'(t+\cdot )$
and $u'_t(\cdot ) = u'(t+\cdot )$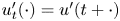 are defined on $[-2\pi,\,0]$
are defined on $[-2\pi,\,0]$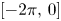 for $t\in [0,\, 2\pi ]$
for $t\in [0,\, 2\pi ]$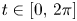 . We completely characterize the well-posedness of ($P_4$
. We completely characterize the well-posedness of ($P_4$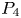 ) in the above two function spaces by using known operator-valued Fourier multiplier theorems.
) in the above two function spaces by using known operator-valued Fourier multiplier theorems.
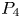
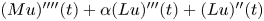
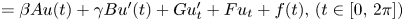
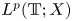
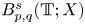
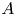
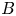
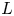
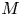
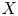
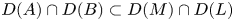

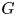
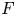
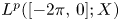
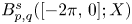
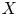
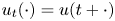
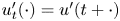
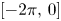
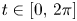
NOVEL STABILITY CONDITIONS FOR SOME GENERALIZATION OF NICHOLSON’S BLOWFLIES MODEL WITH STOCHASTIC PERTURBATIONS
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 64 / Issue 4 / October 2022
- Published online by Cambridge University Press:
- 23 August 2023, pp. 394-405
-
- Article
- Export citation
Spreading dynamics of a diffusive epidemic model with free boundaries and two delays
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 34 / Issue 6 / December 2023
- Published online by Cambridge University Press:
- 11 August 2023, pp. 1133-1169
-
- Article
- Export citation
Local and global existence and uniqueness of solution for abstract differential equations with state-dependent argument
- Part of
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 66 / Issue 2 / May 2023
- Published online by Cambridge University Press:
- 26 April 2023, pp. 305-345
-
- Article
- Export citation
Dynamics of a delayed population patch model with the dispersion matrix incorporating population loss
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 34 / Issue 4 / August 2023
- Published online by Cambridge University Press:
- 21 March 2023, pp. 870-895
-
- Article
- Export citation
HOPF BIFURCATION ANALYSIS OF A FRACTIONAL-ORDER HOLLING–TANNER PREDATOR-PREY MODEL WITH TIME DELAY
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 64 / Issue 1 / January 2022
- Published online by Cambridge University Press:
- 05 April 2022, pp. 23-39
-
- Article
- Export citation
-
We study a fractional-order delayed predator-prey model with Holling–Tanner-type functional response. Mainly, by choosing the delay time
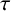 $\tau $ as the bifurcation parameter, we show that Hopf bifurcation can occur as the delay time
$\tau $ as the bifurcation parameter, we show that Hopf bifurcation can occur as the delay time 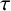 $\tau $ passes some critical values. The local stability of a positive equilibrium and the existence of the Hopf bifurcations are established, and numerical simulations for justifying the theoretical analysis are also presented.
$\tau $ passes some critical values. The local stability of a positive equilibrium and the existence of the Hopf bifurcations are established, and numerical simulations for justifying the theoretical analysis are also presented.
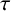
Convergence rate of the EM algorithm for SDEs with low regular drifts
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 59 / Issue 2 / June 2022
- Published online by Cambridge University Press:
- 14 February 2022, pp. 447-470
- Print publication:
- June 2022
-
- Article
- Export citation
On meromorphic solutions of functional-differential equations
- Part of
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 65 / Issue 1 / February 2022
- Published online by Cambridge University Press:
- 10 February 2022, pp. 263-278
-
- Article
- Export citation
-
We consider meromorphic solutions of functional-differential equations
\[ f^{(k)}(z)=a(f^{n}\circ g)(z)+bf(z)+c, \]where $n,\,~k$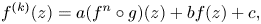
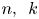 are two positive integers. Firstly, using an elementary method, we describe the forms of $f$
are two positive integers. Firstly, using an elementary method, we describe the forms of $f$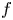 and $g$
and $g$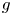 when $f$
when $f$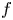 is rational and $a(\neq 0)$
is rational and $a(\neq 0)$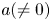 , $b$
, $b$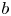 , $c$
, $c$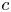 are constants. In addition, by employing Nevanlinna theory, we show that $g$
are constants. In addition, by employing Nevanlinna theory, we show that $g$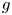 must be linear when $f$
must be linear when $f$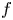 is transcendental and $a(\neq 0)$
is transcendental and $a(\neq 0)$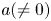 , $b$
, $b$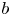 , $c$
, $c$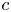 are polynomials in $\mathbb {C}$
are polynomials in $\mathbb {C}$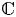 .
.
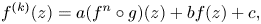
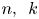
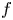
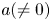
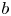
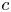
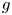
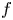
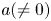
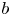
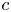
Conley index for manifold-valued retarded functional differential equations without uniqueness of solutions
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 152 / Issue 2 / April 2022
- Published online by Cambridge University Press:
- 19 April 2021, pp. 428-449
- Print publication:
- April 2022
-
- Article
- Export citation
-
We develop a Conley index theory for retarded functional differential equations
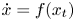 $\dot x=f(x_{t})$ with values in a differentiable manifold and (merely) continuous nonlinearities f. We use this index to establish an existence result for nonconstant full solutions of such equations.
$\dot x=f(x_{t})$ with values in a differentiable manifold and (merely) continuous nonlinearities f. We use this index to establish an existence result for nonconstant full solutions of such equations.
Exact simulation for multivariate Itô diffusions
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 52 / Issue 4 / December 2020
- Published online by Cambridge University Press:
- 03 December 2020, pp. 1003-1034
- Print publication:
- December 2020
-
- Article
- Export citation
