Refine search
Actions for selected content:
9086 results in Discrete Mathematics, Information Theory and Coding
Real-time segmentation and tracking of brain metabolic state in ICU EEG recordings of burst suppression
- from Part II - State space methods for clinical data
-
-
- Book:
- Advanced State Space Methods for Neural and Clinical Data
- Published online:
- 05 October 2015
- Print publication:
- 15 October 2015, pp 330-344
-
- Chapter
- Export citation
Bayesian nonparametric learning of switching dynamics in cohort physiological time series: application in critical care patient monitoring
- from Part II - State space methods for clinical data
-
-
- Book:
- Advanced State Space Methods for Neural and Clinical Data
- Published online:
- 05 October 2015
- Print publication:
- 15 October 2015, pp 257-282
-
- Chapter
- Export citation
Convergence of Achlioptas Processes via Differential Equations with Unique Solutions
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 1 / January 2016
- Published online by Cambridge University Press:
- 14 October 2015, pp. 154-171
-
- Article
- Export citation

Designs and their Codes
-
- Published online:
- 05 October 2015
- Print publication:
- 28 August 1992

Advanced State Space Methods for Neural and Clinical Data
-
- Published online:
- 05 October 2015
- Print publication:
- 15 October 2015
Distinct Distances from Three Points
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 4 / July 2016
- Published online by Cambridge University Press:
- 30 September 2015, pp. 623-632
-
- Article
- Export citation
The Random Graph Intuition for the Tournament Game
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 1 / January 2016
- Published online by Cambridge University Press:
- 23 September 2015, pp. 76-88
-
- Article
- Export citation
Quantitative Equidistribution for Certain Quadruples in Quasi-Random Groups: Erratum
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 3 / May 2016
- Published online by Cambridge University Press:
- 22 September 2015, pp. 484-485
-
- Article
-
- You have access
- Export citation
-
In my recent paper [1] there is a mistake in the proof of Corollary 3. The first line of the displayed equation in that proof asserts that
However, since the paper uses complex-valued representations, the integrand on the right here may not retain the absolute value of that on the left. Without this equality, the proof of Corollary 3 can no longer be reduced to an application of Lemma 2. However, it can be proved directly from Schur Orthogonality along very similar lines to the proof of Lemma 2.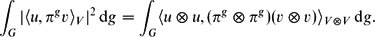 \[\int_G|\langle u,\pi^g v\rangle_V|^2\,\rm{d} g = \int_G\langle u\otimes u,(\pi^g\otimes \pi^g)(v\otimes v)\rangle_{V\otimes V}\, \rm{d} g.\]
\[\int_G|\langle u,\pi^g v\rangle_V|^2\,\rm{d} g = \int_G\langle u\otimes u,(\pi^g\otimes \pi^g)(v\otimes v)\rangle_{V\otimes V}\, \rm{d} g.\]
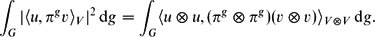
On the Number of Bh-Sets
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 1 / January 2016
- Published online by Cambridge University Press:
- 16 September 2015, pp. 108-129
-
- Article
- Export citation
Sum of Many Dilates
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 3 / May 2016
- Published online by Cambridge University Press:
- 08 September 2015, pp. 460-469
-
- Article
- Export citation
-
We show that for any coprime integers λ1, . . ., λk and any finite A ⊂
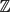 $\mathbb{Z}$ , one haswhere C only depends on λ1, . . ., λk .
$\mathbb{Z}$ , one haswhere C only depends on λ1, . . ., λk .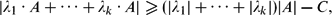 |\lambda_1 \cdot A + \cdots + \lambda_k \cdot A| \geq (|\lambda_1| + \cdots + |\lambda_k|)|A|- C,
|\lambda_1 \cdot A + \cdots + \lambda_k \cdot A| \geq (|\lambda_1| + \cdots + |\lambda_k|)|A|- C,
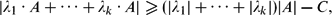
MTK volume 61 Issue 3 Cover and Back matter
-
- Journal:
- Mathematika / Volume 61 / Issue 3 / September 2015
- Published online by Cambridge University Press:
- 16 September 2015, pp. b1-b3
- Print publication:
- September 2015
-
- Article
-
- You have access
- Export citation
MTK volume 61 Issue 3 Cover and Front matter
-
- Journal:
- Mathematika / Volume 61 / Issue 3 / September 2015
- Published online by Cambridge University Press:
- 16 September 2015, pp. f1-f2
- Print publication:
- September 2015
-
- Article
-
- You have access
- Export citation
On the Method of Typical Bounded Differences
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 2 / March 2016
- Published online by Cambridge University Press:
- 27 August 2015, pp. 269-299
-
- Article
- Export citation
A Stability Result for the Union-Closed Size Problem
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 3 / May 2016
- Published online by Cambridge University Press:
- 18 August 2015, pp. 399-418
-
- Article
- Export citation
-
A family of sets is called union-closed if whenever A and B are sets of the family, so is A ∪ B. The long-standing union-closed conjecture states that if a family of subsets of [n] is union-closed, some element appears in at least half the sets of the family. A natural weakening is that the union-closed conjecture holds for large families, that is, families consisting of at least p 02n sets for some constant p 0. The first result in this direction appears in a recent paper of Balla, Bollobás and Eccles [1], who showed that union-closed families of at least
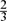 $\tfrac{2}{3}$ 2n sets satisfy the conjecture; they proved this by determining the minimum possible average size of a set in a union-closed family of given size. However, the methods used in that paper cannot prove a better constant than $\tfrac{2}{3}$ . Here, we provide a stability result for the main theorem of [1], and as a consequence we prove the union-closed conjecture for families of at least ( $\tfrac{2}{3}$ − c)2n sets, for a positive constant c.
$\tfrac{2}{3}$ 2n sets satisfy the conjecture; they proved this by determining the minimum possible average size of a set in a union-closed family of given size. However, the methods used in that paper cannot prove a better constant than $\tfrac{2}{3}$ . Here, we provide a stability result for the main theorem of [1], and as a consequence we prove the union-closed conjecture for families of at least ( $\tfrac{2}{3}$ − c)2n sets, for a positive constant c.
Simple Containers for Simple Hypergraphs
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 3 / May 2016
- Published online by Cambridge University Press:
- 17 August 2015, pp. 448-459
-
- Article
- Export citation

Inequalities for Graph Eigenvalues
-
- Published online:
- 05 August 2015
- Print publication:
- 23 July 2015
The Satisfiability Threshold for k-XORSAT
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 2 / March 2016
- Published online by Cambridge University Press:
- 31 July 2015, pp. 236-268
-
- Article
- Export citation
-
We consider ‘unconstrained’ random k-XORSAT, which is a uniformly random system of m linear non-homogeneous equations in
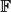 $\mathbb{F}$ 2 over n variables, each equation containing k ⩾ 3 variables, and also consider a ‘constrained’ model where every variable appears in at least two equations. Dubois and Mandler proved that m/n = 1 is a sharp threshold for satisfiability of constrained 3-XORSAT, and analysed the 2-core of a random 3-uniform hypergraph to extend this result to find the threshold for unconstrained 3-XORSAT.
$\mathbb{F}$ 2 over n variables, each equation containing k ⩾ 3 variables, and also consider a ‘constrained’ model where every variable appears in at least two equations. Dubois and Mandler proved that m/n = 1 is a sharp threshold for satisfiability of constrained 3-XORSAT, and analysed the 2-core of a random 3-uniform hypergraph to extend this result to find the threshold for unconstrained 3-XORSAT.We show that m/n = 1 remains a sharp threshold for satisfiability of constrained k-XORSAT for every k ⩾ 3, and we use standard results on the 2-core of a random k-uniform hypergraph to extend this result to find the threshold for unconstrained k-XORSAT. For constrained k-XORSAT we narrow the phase transition window, showing that m − n → −∞ implies almost-sure satisfiability, while m − n → +∞ implies almost-sure unsatisfiability.
Universality of Graphs with Few Triangles and Anti-Triangles
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 4 / July 2016
- Published online by Cambridge University Press:
- 29 July 2015, pp. 560-576
-
- Article
- Export citation
-
We study 3-random-like graphs, that is, sequences of graphs in which the densities of triangles and anti-triangles converge to 1/8. Since the random graph
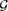 $\mathcal{G}$ n,1/2 is, in particular, 3-random-like, this can be viewed as a weak version of quasi-randomness. We first show that 3-random-like graphs are 4-universal, that is, they contain induced copies of all 4-vertex graphs. This settles a question of Linial and Morgenstern [10]. We then show that for larger subgraphs, 3-random-like sequences demonstrate completely different behaviour. We prove that for every graph H on n ⩾ 13 vertices there exist 3-random-like graphs without an induced copy of H. Moreover, we prove that for every ℓ there are 3-random-like graphs which are ℓ-universal but not m-universal when m is sufficiently large compared to ℓ.
$\mathcal{G}$ n,1/2 is, in particular, 3-random-like, this can be viewed as a weak version of quasi-randomness. We first show that 3-random-like graphs are 4-universal, that is, they contain induced copies of all 4-vertex graphs. This settles a question of Linial and Morgenstern [10]. We then show that for larger subgraphs, 3-random-like sequences demonstrate completely different behaviour. We prove that for every graph H on n ⩾ 13 vertices there exist 3-random-like graphs without an induced copy of H. Moreover, we prove that for every ℓ there are 3-random-like graphs which are ℓ-universal but not m-universal when m is sufficiently large compared to ℓ.
Contents
-
- Book:
- Inequalities for Graph Eigenvalues
- Published online:
- 05 August 2015
- Print publication:
- 23 July 2015, pp v-viii
-
- Chapter
- Export citation
6 - Laplacian eigenvalues
-
- Book:
- Inequalities for Graph Eigenvalues
- Published online:
- 05 August 2015
- Print publication:
- 23 July 2015, pp 155-203
-
- Chapter
- Export citation
