Let {Xn, n = 0, 1, 2, ···} be a transient Markov chain which, when restricted to the state space 𝒩 + = {1, 2, ···}, is governed by an irreducible, aperiodic and strictly substochastic matrix 𝐏 = (pij), and let pij(n) = P ∈ Xn = j, Xk ∈ 𝒩+ for k = 0, 1, ···, n | X0 = i], i, j 𝒩 +. The prime concern of this paper is conditions for the existence of the limits, qij say, of 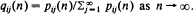 as n →∞. If
as n →∞. If 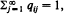 the distribution (qij) is called the quasi-stationary distribution of {Xn} and has considerable practical importance. It will be shown that, under some conditions, if a non-negative non-trivial vector x = (xi) satisfying rxT = xT𝐏 and
the distribution (qij) is called the quasi-stationary distribution of {Xn} and has considerable practical importance. It will be shown that, under some conditions, if a non-negative non-trivial vector x = (xi) satisfying rxT = xT𝐏 and 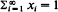 exists, where r is the convergence norm of 𝐏, i.e. r = R–1 and
exists, where r is the convergence norm of 𝐏, i.e. r = R–1 and 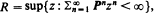 and T denotes transpose, then it is unique, positive elementwise, and qij(n) necessarily converge to xj as n →∞. Unlike existing results in the literature, our results can be applied even to the R-null and R-transient cases. Finally, an application to a left-continuous random walk whose governing substochastic matrix is R-transient is discussed to demonstrate the usefulness of our results.
and T denotes transpose, then it is unique, positive elementwise, and qij(n) necessarily converge to xj as n →∞. Unlike existing results in the literature, our results can be applied even to the R-null and R-transient cases. Finally, an application to a left-continuous random walk whose governing substochastic matrix is R-transient is discussed to demonstrate the usefulness of our results.
Let P be an irreducible Markov matrix. Assume:

We study conditions for such a matrix to be nonrecurrent. If P is nonrecurrent we study the invariant vectors of P (invariant column vectors and invariant row vectors).
In this paper X(t) denotes Brownian motion on the line 0 ≤ t < ∞, E is a compact subset of (0, ∞) and F a compact subset of ( -∞, ∞). Then δ(E, F) is the supremum of the numbers c such that
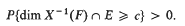
In [4, 5] some lower and upper bounds for δ were found in terms of dim E and dim F, and it seemed possible that δ(E, F) could be determined entirely by these constants; this much is false, as the examples in our last paragraph will demonstrate. Here we shall show that δ(E, F) depends on a certain metric character η(E × F). However, η is not calculated relative to the Euclidean metric: the set F must be compressed to compensate for the oscillations of most paths X(t). Fortunately, η(E × F) can be calculated for a large enough class of sets E and F, by means of sequences of integers, to test any conjectures (and disprove most of them). In passing from η to δ we present a slight variation of Frostman's theory [3II].

