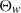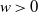Refine listing
Actions for selected content:
1723 results in 60Jxx
Perturbation theory for killed Markov processes and quasi-stationary distributions
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 19 March 2025, pp. 1138-1165
- Print publication:
- September 2025
-
- Article
- Export citation
-
Motivated by recent developments of quasi-stationary Monte Carlo methods, we investigate the stability of quasi-stationary distributions of killed Markov processes under perturbations of the generator. We first consider a general bounded self-adjoint perturbation operator, and then study a particular unbounded perturbation corresponding to truncation of the killing rate. In both scenarios, we quantify the difference between eigenfunctions of the smallest eigenvalue of the perturbed and unperturbed generators in a Hilbert space norm. As a consequence,
 $\mathcal{L}^1$-norm estimates of the difference of the resulting quasi-stationary distributions in terms of the perturbation are provided.
$\mathcal{L}^1$-norm estimates of the difference of the resulting quasi-stationary distributions in terms of the perturbation are provided.

The speed of a biased walk on a Galton–Watson tree without leaves is monotonic for low values of bias
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 05 March 2025, pp. 1044-1052
- Print publication:
- September 2025
-
- Article
- Export citation
-
We show that for
 $\lambda\in[0,{m_1}/({1+\sqrt{1-{1}/{m_1}}})]$, the biased random walk’s speed on a Galton–Watson tree without leaves is strictly decreasing, where
$\lambda\in[0,{m_1}/({1+\sqrt{1-{1}/{m_1}}})]$, the biased random walk’s speed on a Galton–Watson tree without leaves is strictly decreasing, where  $m_1\geq 2$. Our result extends the monotonic interval of the speed on a Galton–Watson tree.
$m_1\geq 2$. Our result extends the monotonic interval of the speed on a Galton–Watson tree.


Phase-type representations for exponential distributions
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 04 March 2025, pp. 969-999
- Print publication:
- September 2025
-
- Article
- Export citation
A non-convex optimization approach of searching algebraic degree phase-type representations for general phase-type distributions
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 03 March 2025, pp. 1101-1137
- Print publication:
- September 2025
-
- Article
- Export citation
Long edges in Galton–Watson trees
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 18 February 2025, pp. 1053-1068
- Print publication:
- September 2025
-
- Article
- Export citation
Optimal convergence rates of MCMC integration for functions with unbounded second moment
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 14 February 2025, pp. 1069-1075
- Print publication:
- September 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We study the Markov chain Monte Carlo estimator for numerical integration for functions that do not need to be square integrable with respect to the invariant distribution. For chains with a spectral gap we show that the absolute mean error for
 $L^p$ functions, with
$L^p$ functions, with  $p \in (1,2)$, decreases like
$p \in (1,2)$, decreases like  $n^{({1}/{p}) -1}$, which is known to be the optimal rate. This improves currently known results where an additional parameter
$n^{({1}/{p}) -1}$, which is known to be the optimal rate. This improves currently known results where an additional parameter  $\delta \gt 0$ appears and the convergence is of order
$\delta \gt 0$ appears and the convergence is of order  $n^{(({1+\delta})/{p})-1}$.
$n^{(({1+\delta})/{p})-1}$.





Critical configurations of the hard-core model on square grid graphs
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 34 / Issue 3 / May 2025
- Published online by Cambridge University Press:
- 04 February 2025, pp. 445-485
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider the hard-core model on a finite square grid graph with stochastic Glauber dynamics parametrized by the inverse temperature
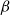 $\beta$. We investigate how the transition between its two maximum-occupancy configurations takes place in the low-temperature regime
$\beta$. We investigate how the transition between its two maximum-occupancy configurations takes place in the low-temperature regime 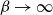 $\beta \to \infty$ in the case of periodic boundary conditions. The hard-core constraints and the grid symmetry make the structure of the critical configurations for this transition, also known as essential saddles, very rich and complex. We provide a comprehensive geometrical characterization of these configurations that together constitute a bottleneck for the Glauber dynamics in the low-temperature limit. In particular, we develop a novel isoperimetric inequality for hard-core configurations with a fixed number of particles and show how the essential saddles are characterized not only by the number of particles but also their geometry.
$\beta \to \infty$ in the case of periodic boundary conditions. The hard-core constraints and the grid symmetry make the structure of the critical configurations for this transition, also known as essential saddles, very rich and complex. We provide a comprehensive geometrical characterization of these configurations that together constitute a bottleneck for the Glauber dynamics in the low-temperature limit. In particular, we develop a novel isoperimetric inequality for hard-core configurations with a fixed number of particles and show how the essential saddles are characterized not only by the number of particles but also their geometry.
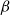
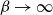
The number of individuals alive in a branching process given only times of deaths
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 31 January 2025, pp. 871-906
- Print publication:
- September 2025
-
- Article
- Export citation
Asymptotic expansion of the invariant measure for Markov-modulated ODEs at high frequency
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 30 January 2025, pp. 898-924
- Print publication:
- September 2025
-
- Article
- Export citation
-
We consider time-inhomogeneous ordinary differential equations (ODEs) whose parameters are governed by an underlying ergodic Markov process. When this underlying process is accelerated by a factor
 $\varepsilon^{-1}$, an averaging phenomenon occurs and the solution of the ODE converges to a deterministic ODE as
$\varepsilon^{-1}$, an averaging phenomenon occurs and the solution of the ODE converges to a deterministic ODE as  $\varepsilon$ vanishes. We are interested in cases where this averaged flow is globally attracted to a point. In that case, the equilibrium distribution of the solution of the ODE converges to a Dirac mass at this point. We prove an asymptotic expansion in terms of
$\varepsilon$ vanishes. We are interested in cases where this averaged flow is globally attracted to a point. In that case, the equilibrium distribution of the solution of the ODE converges to a Dirac mass at this point. We prove an asymptotic expansion in terms of  $\varepsilon$ for this convergence, with a somewhat explicit formula for the first-order term. The results are applied in three contexts: linear Markov-modulated ODEs, randomized splitting schemes, and Lotka–Volterra models in a random environment. In particular, as a corollary, we prove the existence of two matrices whose convex combinations are all stable but are such that, for a suitable jump rate, the top Lyapunov exponent of a Markov-modulated linear ODE switching between these two matrices is positive.
$\varepsilon$ for this convergence, with a somewhat explicit formula for the first-order term. The results are applied in three contexts: linear Markov-modulated ODEs, randomized splitting schemes, and Lotka–Volterra models in a random environment. In particular, as a corollary, we prove the existence of two matrices whose convex combinations are all stable but are such that, for a suitable jump rate, the top Lyapunov exponent of a Markov-modulated linear ODE switching between these two matrices is positive.



Perturbation analysis of Markov chain Monte Carlo for graphical models
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 06 January 2025, pp. 950-971
- Print publication:
- September 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Mean-field analysis of stochastic networks with reservation
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 2 / June 2025
- Published online by Cambridge University Press:
- 06 January 2025, pp. 832-858
- Print publication:
- June 2025
-
- Article
- Export citation
-
The problem of reservation in a large distributed system is analyzed via a new mathematical model. The target application is car-sharing systems. This model is motivated by the large station-based car-sharing system in France called Autolib’. This system can be described as a closed stochastic network where the nodes are the stations and the customers are the cars. The user can reserve a car and a parking space. We study the evolution of the system when the reservation of parking spaces and cars is effective for all users. The asymptotic behavior of the underlying stochastic network is given when the number N of stations and the fleet size M increase at the same rate. The analysis involves a Markov process on a state space with dimension of order
 $N^2$. It is quite remarkable that the state process describing the evolution of the stations, whose dimension is of order N, converges in distribution, although not Markov, to a non-homogeneous Markov process. We prove this mean-field convergence. We also prove, using combinatorial arguments, that the mean-field limit has a unique equilibrium measure when the time between reserving and picking up the car is sufficiently small. This result extends the case where only the parking space can be reserved.
$N^2$. It is quite remarkable that the state process describing the evolution of the stations, whose dimension is of order N, converges in distribution, although not Markov, to a non-homogeneous Markov process. We prove this mean-field convergence. We also prove, using combinatorial arguments, that the mean-field limit has a unique equilibrium measure when the time between reserving and picking up the car is sufficiently small. This result extends the case where only the parking space can be reserved.

Fractional Brownian motion with deterministic drift: how critical is drift regularity in hitting probabilities
- Part of
-
- Journal:
- Mathematical Proceedings of the Cambridge Philosophical Society / Volume 178 / Issue 1 / January 2025
- Published online by Cambridge University Press:
- 20 February 2025, pp. 103-132
- Print publication:
- January 2025
-
- Article
- Export citation
-
Let
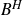 $B^{H}$ be a d-dimensional fractional Brownian motion with Hurst index
$B^{H}$ be a d-dimensional fractional Brownian motion with Hurst index 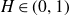 $H\in(0,1)$,
$H\in(0,1)$, 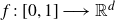 $f\,:\,[0,1]\longrightarrow\mathbb{R}^{d}$ a Borel function, and
$f\,:\,[0,1]\longrightarrow\mathbb{R}^{d}$ a Borel function, and 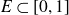 $E\subset[0,1]$,
$E\subset[0,1]$, 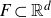 $F\subset\mathbb{R}^{d}$ are given Borel sets. The focus of this paper is on hitting probabilities of the non-centered Gaussian process
$F\subset\mathbb{R}^{d}$ are given Borel sets. The focus of this paper is on hitting probabilities of the non-centered Gaussian process 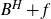 $B^{H}+f$. It aims to highlight how each component f, E and F is involved in determining the upper and lower bounds of
$B^{H}+f$. It aims to highlight how each component f, E and F is involved in determining the upper and lower bounds of 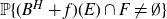 $\mathbb{P}\{(B^H+f)(E)\cap F\neq \emptyset \}$. When F is a singleton and f is a general measurable drift, some new estimates are obtained for the last probability by means of suitable Hausdorff measure and capacity of the graph
$\mathbb{P}\{(B^H+f)(E)\cap F\neq \emptyset \}$. When F is a singleton and f is a general measurable drift, some new estimates are obtained for the last probability by means of suitable Hausdorff measure and capacity of the graph 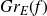 $Gr_E(f)$. As application we deal with the issue of polarity of points for
$Gr_E(f)$. As application we deal with the issue of polarity of points for 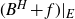 $(B^H+f)\vert_E$ (the restriction of
$(B^H+f)\vert_E$ (the restriction of 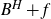 $B^H+f$ to the subset
$B^H+f$ to the subset 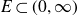 $E\subset (0,\infty)$).
$E\subset (0,\infty)$).
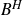
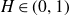
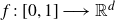
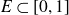
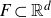
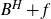
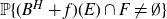
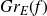
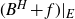
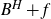
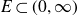
The Markov chain embedding problem in a one-jump setting
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 2 / June 2025
- Published online by Cambridge University Press:
- 09 December 2024, pp. 674-696
- Print publication:
- June 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The embedding problem of Markov chains examines whether a stochastic matrix
 $\mathbf{P} $ can arise as the transition matrix from time 0 to time 1 of a continuous-time Markov chain. When the chain is homogeneous, it checks if
$\mathbf{P} $ can arise as the transition matrix from time 0 to time 1 of a continuous-time Markov chain. When the chain is homogeneous, it checks if  $ \mathbf{P}=\exp{\mathbf{Q}}$ for a rate matrix
$ \mathbf{P}=\exp{\mathbf{Q}}$ for a rate matrix  $ \mathbf{Q}$ with zero row sums and non-negative off-diagonal elements, called a Markov generator. It is known that a Markov generator may not always exist or be unique. This paper addresses finding
$ \mathbf{Q}$ with zero row sums and non-negative off-diagonal elements, called a Markov generator. It is known that a Markov generator may not always exist or be unique. This paper addresses finding  $ \mathbf{Q}$, assuming that the process has at most one jump per unit time interval, and focuses on the problem of aligning the conditional one-jump transition matrix from time 0 to time 1 with
$ \mathbf{Q}$, assuming that the process has at most one jump per unit time interval, and focuses on the problem of aligning the conditional one-jump transition matrix from time 0 to time 1 with  $ \mathbf{P}$. We derive a formula for this matrix in terms of
$ \mathbf{P}$. We derive a formula for this matrix in terms of  $ \mathbf{Q}$ and establish that for any
$ \mathbf{Q}$ and establish that for any  $ \mathbf{P}$ with non-zero diagonal entries, a unique
$ \mathbf{P}$ with non-zero diagonal entries, a unique  $ \mathbf{Q}$, called the
$ \mathbf{Q}$, called the  ${\unicode{x1D7D9}}$-generator, exists. We compare the
${\unicode{x1D7D9}}$-generator, exists. We compare the  ${\unicode{x1D7D9}}$-generator with the one-jump rate matrix from Jarrow, Lando, and Turnbull (1997), showing which is a better approximate Markov generator of
${\unicode{x1D7D9}}$-generator with the one-jump rate matrix from Jarrow, Lando, and Turnbull (1997), showing which is a better approximate Markov generator of  $ \mathbf{P}$ in some practical cases.
$ \mathbf{P}$ in some practical cases.











Sampling from the random cluster model on random regular graphs at all temperatures via Glauber dynamics
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 34 / Issue 3 / May 2025
- Published online by Cambridge University Press:
- 09 December 2024, pp. 359-391
-
- Article
- Export citation
-
We consider the performance of Glauber dynamics for the random cluster model with real parameter
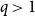 $q\gt 1$ and temperature
$q\gt 1$ and temperature 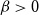 $\beta \gt 0$. Recent work by Helmuth, Jenssen, and Perkins detailed the ordered/disordered transition of the model on random
$\beta \gt 0$. Recent work by Helmuth, Jenssen, and Perkins detailed the ordered/disordered transition of the model on random 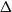 $\Delta$-regular graphs for all sufficiently large
$\Delta$-regular graphs for all sufficiently large 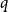 $q$ and obtained an efficient sampling algorithm for all temperatures
$q$ and obtained an efficient sampling algorithm for all temperatures 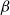 $\beta$ using cluster expansion methods. Despite this major progress, the performance of natural Markov chains, including Glauber dynamics, is not yet well understood on the random regular graph, partly because of the non-local nature of the model (especially at low temperatures) and partly because of severe bottleneck phenomena that emerge in a window around the ordered/disordered transition. Nevertheless, it is widely conjectured that the bottleneck phenomena that impede mixing from worst-case starting configurations can be avoided by initialising the chain more judiciously. Our main result establishes this conjecture for all sufficiently large
$\beta$ using cluster expansion methods. Despite this major progress, the performance of natural Markov chains, including Glauber dynamics, is not yet well understood on the random regular graph, partly because of the non-local nature of the model (especially at low temperatures) and partly because of severe bottleneck phenomena that emerge in a window around the ordered/disordered transition. Nevertheless, it is widely conjectured that the bottleneck phenomena that impede mixing from worst-case starting configurations can be avoided by initialising the chain more judiciously. Our main result establishes this conjecture for all sufficiently large 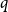 $q$ (with respect to
$q$ (with respect to 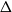 $\Delta$). Specifically, we consider the mixing time of Glauber dynamics initialised from the two extreme configurations, the all-in and all-out, and obtain a pair of fast mixing bounds which cover all temperatures
$\Delta$). Specifically, we consider the mixing time of Glauber dynamics initialised from the two extreme configurations, the all-in and all-out, and obtain a pair of fast mixing bounds which cover all temperatures 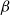 $\beta$, including in particular the bottleneck window. Our result is inspired by the recent approach of Gheissari and Sinclair for the Ising model who obtained a similar flavoured mixing-time bound on the random regular graph for sufficiently low temperatures. To cover all temperatures in the RC model, we refine appropriately the structural results of Helmuth, Jenssen and Perkins about the ordered/disordered transition and show spatial mixing properties ‘within the phase’, which are then related to the evolution of the chain.
$\beta$, including in particular the bottleneck window. Our result is inspired by the recent approach of Gheissari and Sinclair for the Ising model who obtained a similar flavoured mixing-time bound on the random regular graph for sufficiently low temperatures. To cover all temperatures in the RC model, we refine appropriately the structural results of Helmuth, Jenssen and Perkins about the ordered/disordered transition and show spatial mixing properties ‘within the phase’, which are then related to the evolution of the chain.
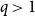
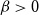
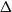
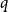
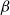
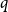
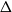
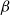
Optimal stopping of Gauss–Markov bridges
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 1 / March 2025
- Published online by Cambridge University Press:
- 04 December 2024, pp. 1-34
- Print publication:
- March 2025
-
- Article
- Export citation
Discounted optimal stopping zero-sum games in diffusion type models with maxima and minima
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 1 / March 2025
- Published online by Cambridge University Press:
- 03 December 2024, pp. 241-270
- Print publication:
- March 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
On the Brownian range and the Brownian reversal
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 2 / June 2025
- Published online by Cambridge University Press:
- 03 December 2024, pp. 775-794
- Print publication:
- June 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Convergence of the derivative martingale for the branching random walk in time-inhomogeneous random environment
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 2 / June 2025
- Published online by Cambridge University Press:
- 02 December 2024, pp. 642-676
- Print publication:
- June 2025
-
- Article
- Export citation
Gaussian fluctuations for the two-urn model
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 2 / June 2025
- Published online by Cambridge University Press:
- 02 December 2024, pp. 607-641
- Print publication:
- June 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We introduce a modification of the generalized Pólya urn model containing two urns, and we study the number of balls
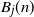 $B_j(n)$ of a given color
$B_j(n)$ of a given color 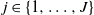 $j\in\{1,\ldots,J\}$ added to the urns after n draws, where
$j\in\{1,\ldots,J\}$ added to the urns after n draws, where 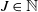 $J\in\mathbb{N}$. We provide sufficient conditions under which the random variables
$J\in\mathbb{N}$. We provide sufficient conditions under which the random variables 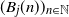 $(B_j(n))_{n\in\mathbb{N}}$, properly normalized and centered, converge weakly to a limiting random variable. The result reveals a similar trichotomy as in the classical case with one urn, one of the main differences being that in the scaling we encounter 1-periodic continuous functions. Another difference in our results compared to the classical urn models is that the phase transition of the second-order behavior occurs at
$(B_j(n))_{n\in\mathbb{N}}$, properly normalized and centered, converge weakly to a limiting random variable. The result reveals a similar trichotomy as in the classical case with one urn, one of the main differences being that in the scaling we encounter 1-periodic continuous functions. Another difference in our results compared to the classical urn models is that the phase transition of the second-order behavior occurs at 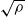 $\sqrt{\rho}$ and not at
$\sqrt{\rho}$ and not at 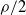 $\rho/2$, where
$\rho/2$, where 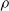 $\rho$ is the dominant eigenvalue of the mean replacement matrix.
$\rho$ is the dominant eigenvalue of the mean replacement matrix.
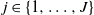
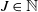
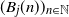
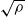
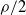
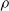
Some results on the supremum and on the first passage time of the generalized telegraph process
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 2 / June 2025
- Published online by Cambridge University Press:
- 02 December 2024, pp. 453-491
- Print publication:
- June 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We analyze the process M(t) representing the maximum of the one-dimensional telegraph process X(t) with exponentially distributed upward random times and generally distributed downward random times. The evolution of M(t) is governed by an alternating renewal of two phases: a rising phase R and a constant phase C. During a rising phase, X(t) moves upward, whereas, during a constant phase, it moves upward and downward, continuing to move until it attains the maximal level previously reached. Under some choices of the distribution of the downward times, we are able to determine the distribution of C, which allows us to obtain some bounds for the survival function of M(t). In the particular case of exponential downward random times, we derive an explicit expression for the survival function of M(t). Finally, the moments of the first passage time
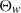 $\Theta_w$ of the process X(t) through a fixed boundary
$\Theta_w$ of the process X(t) through a fixed boundary 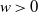 $w>0$ are analyzed.
$w>0$ are analyzed.