A model is proposed for the one-dimensional spectrum and streamwise Reynolds stress in pipe flow for arbitrarily large Reynolds numbers. Constructed in wavenumber space, the model comprises four principal contributions to the spectrum: streaks, large-scale motions, very-large-scale motions and incoherent turbulence. It accounts for the broad and overlapping spectral content of these contributions from different eddy types. The model reproduces well the broad structure of the premultiplied one-dimensional spectrum of the streamwise velocity, although the bimodal shape that has been observed at certain wall-normal locations, and the  $-5/3$ slope of the inertial subrange, are not captured effectively because of the simplifications made within the model. Regardless, the Reynolds stress distribution is well reproduced, even within the near-wall region, including key features of wall-bounded flows such as the Reynolds number dependence of the inner peak, the formation of a logarithmic region, and the formation of an outer peak. These findings suggest that many of these features arise from the overlap of energy content produced by both inner- and outer-scaled eddy structures combined with the viscous-scaled influence of the wall. The model is also used to compare with canonical turbulent boundary layer and channel flows, and despite some differences being apparent, we speculate that with only minor modifications to its coefficients, the model can be adapted to these flows as well.
$-5/3$ slope of the inertial subrange, are not captured effectively because of the simplifications made within the model. Regardless, the Reynolds stress distribution is well reproduced, even within the near-wall region, including key features of wall-bounded flows such as the Reynolds number dependence of the inner peak, the formation of a logarithmic region, and the formation of an outer peak. These findings suggest that many of these features arise from the overlap of energy content produced by both inner- and outer-scaled eddy structures combined with the viscous-scaled influence of the wall. The model is also used to compare with canonical turbulent boundary layer and channel flows, and despite some differences being apparent, we speculate that with only minor modifications to its coefficients, the model can be adapted to these flows as well.
Direct numerical simulations are performed for turbulent forced convection in a half-channel flow with a wall oscillating either as a spanwise plane oscillation or to generate a streamwise travelling wave. The friction Reynolds number is fixed at  $Re_{\tau _0} = 590$, but the Prandtl number
$Re_{\tau _0} = 590$, but the Prandtl number  $Pr$ is varied from 0.71 to 20. For
$Pr$ is varied from 0.71 to 20. For  $Pr\gt 1$, the heat transfer is reduced by more than the drag, 40 % compared with 30 % at
$Pr\gt 1$, the heat transfer is reduced by more than the drag, 40 % compared with 30 % at  $Pr=7.5$. This outcome is related to the different responses of the velocity and thermal fields to the Stokes layer. It is shown that the Stokes layer near the wall attenuates the large-scale energy of the turbulent heat flux and the turbulent shear stress, but amplifies their small-scale energy. At higher Prandtl numbers, the thinning of the conductive sublayer means that the energetic scales of the turbulent heat flux move closer to the wall, where they are exposed to a stronger Stokes layer production, increasing the contribution of the small-scale energy amplification. A predictive model is derived for the Reynolds and Prandtl number dependence of the heat-transfer reduction based on the scaling of the thermal statistics. The model agrees well with the computations for Prandtl numbers up to 20.
$Pr=7.5$. This outcome is related to the different responses of the velocity and thermal fields to the Stokes layer. It is shown that the Stokes layer near the wall attenuates the large-scale energy of the turbulent heat flux and the turbulent shear stress, but amplifies their small-scale energy. At higher Prandtl numbers, the thinning of the conductive sublayer means that the energetic scales of the turbulent heat flux move closer to the wall, where they are exposed to a stronger Stokes layer production, increasing the contribution of the small-scale energy amplification. A predictive model is derived for the Reynolds and Prandtl number dependence of the heat-transfer reduction based on the scaling of the thermal statistics. The model agrees well with the computations for Prandtl numbers up to 20.
A new wall-wake law is proposed for the streamwise turbulence in the outer region of a turbulent boundary layer. The formulation pairs the logarithmic part of the profile (with a slope  $A_1$ and additive constant
$A_1$ and additive constant  $B_1$) to an outer linear part, and it accurately describes over 95 % of the boundary layer profile at high Reynolds numbers. Once the slope
$B_1$) to an outer linear part, and it accurately describes over 95 % of the boundary layer profile at high Reynolds numbers. Once the slope  $A_1$ is fixed,
$A_1$ is fixed,  $B_1$ is the only free parameter determining the fit. Most importantly,
$B_1$ is the only free parameter determining the fit. Most importantly,  $B_1$ is shown to follow the same trend with Reynolds number as the wake factor in the wall-wake law for the mean velocity, which is tied to changes in scaling of the mean flow and the turbulence that occur at low Reynolds number.
$B_1$ is shown to follow the same trend with Reynolds number as the wake factor in the wall-wake law for the mean velocity, which is tied to changes in scaling of the mean flow and the turbulence that occur at low Reynolds number.
The wall dependence of length scales used to describe large- and small-scale structures of turbulence is examined using highly resolved experiments in zero-pressure-gradient turbulent boundary layers and pipe flows spanning the range 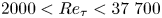 $2000< Re_\tau <37\ 700$. Of particular interest is the influence of external intermittency on the scaling of these length scales. It is found that when suitable scaling parameters are selected and external intermittency is accounted for, the dissipative motions follow inner scaling even into the outer-scaled regions of the flow, and that certain large-scale descriptions follow outer scaling even in the inner-scaled regions of the flow. The wall dependence is the same for both internal pipe and external boundary layer flows, and the different length scales can be related to recognizable features in the longitudinal wavenumber spectrum.
$2000< Re_\tau <37\ 700$. Of particular interest is the influence of external intermittency on the scaling of these length scales. It is found that when suitable scaling parameters are selected and external intermittency is accounted for, the dissipative motions follow inner scaling even into the outer-scaled regions of the flow, and that certain large-scale descriptions follow outer scaling even in the inner-scaled regions of the flow. The wall dependence is the same for both internal pipe and external boundary layer flows, and the different length scales can be related to recognizable features in the longitudinal wavenumber spectrum.
Vortical impulse theory is used to investigate the relationship between turbine thrust and the near-wake velocity and vorticity fields. Three different hypotheses regarding the near-wake structure allow the derivation of novel expressions for the thrust on a steadily rotating wind turbine, and these are tested using stereoscopic particle-image velocimetry (PIV) data acquired just behind a rotor in a water channel. When one assumes that vortex lines and streamlines are aligned in a rotor-fixed frame of reference, one obtains a PIV-based thrust estimate that fails even to capture the trend of the directly measured thrust, and this failure is attributed to an implicit assumption that most of the generated thrust does useful work. When one neglects the axial gradients of radial velocity, the PIV-based thrust estimate captures the measured thrust trend, but underpredicts its magnitude by approximately 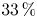 $33\,\%$. The third and most promising physical proposition treats the trailing vortices as purely ‘rolling’ structures that exhibit zero-strain rate in their cores, with the corresponding thrust estimates in close agreement with direct thrust measurements. This best-performing expression appears as a correction to the classical thrust expression from momentum theory, possessing additional squared-velocity terms that can account for the high-thrust regime of turbine operation that is typically addressed empirically.
$33\,\%$. The third and most promising physical proposition treats the trailing vortices as purely ‘rolling’ structures that exhibit zero-strain rate in their cores, with the corresponding thrust estimates in close agreement with direct thrust measurements. This best-performing expression appears as a correction to the classical thrust expression from momentum theory, possessing additional squared-velocity terms that can account for the high-thrust regime of turbine operation that is typically addressed empirically.

