Refine search
Actions for selected content:
104 results
Propagation dynamics of time-dependent reaction-diffusion equations under climate change in an infinite cylinder
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 19 December 2025, pp. 1-22
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In this paper, we consider a reaction-diffusion equation that models the time-almost periodic response to climate change within a straight, infinite cylindrical domain. The shifting edge of the habitat is characterised by a time-almost periodic function, reflecting the varying pace of environmental changes. Note that the principal spectral theory is an important role to study the dynamics of reaction-diffusion equations in time heterogeneous environment. Initially, for time-almost periodic parabolic equations in finite cylindrical domains, we develop the principal spectral theory of such equations with mixed Dirichlet–Neumann boundary conditions. Subsequently, we demonstrate that the approximate principal Lyapunov exponent serves as a definitive threshold for species persistence versus extinction. Then, the existence, exponential decay and stability of the forced wave solutions
 $U(t,x_{1},y)=V\left (t,x_{1}-\int ^{t}_{0}c(s)ds,y\right )$ are established. Additionally, we analyse how fluctuations in the shifting speed affect the approximate top Lyapunov exponent.
$U(t,x_{1},y)=V\left (t,x_{1}-\int ^{t}_{0}c(s)ds,y\right )$ are established. Additionally, we analyse how fluctuations in the shifting speed affect the approximate top Lyapunov exponent.

Impact of birth pulse and environment shift on population survival and propagation
- Part of
-
- Journal:
- Canadian Journal of Mathematics , First View
- Published online by Cambridge University Press:
- 17 October 2025, pp. 1-27
-
- Article
- Export citation
Global dynamics of a degenerate reaction-diffusion cholera model with phage-bacteria interaction in a heterogeneous environment
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 22 September 2025, pp. 1-32
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
To investigate multiple effects of the interaction between V. cholerae and phage on cholera transmission, we propose a degenerate reaction-diffusion model with different dispersal rates, which incorporates a short-lived hyperinfectious (HI vibrios) state of V. cholerae and lower-infectious (LI vibrios) state of V. cholerae. Our main purpose is to investigate the existence and stability analysis of multi-class boundary steady states, which is much more complicated and challenging than the case when the boundary steady state is unique. In a spatially heterogeneous case, the basic reproduction number
 $\mathscr{R}_{0}$ is defined as the spectral radius of the sum of two linear operators associated with HI vibrios infection and LI vibrios infection. If
$\mathscr{R}_{0}$ is defined as the spectral radius of the sum of two linear operators associated with HI vibrios infection and LI vibrios infection. If  $\mathscr{R}_{0}\leq 1$, the disease-free steady state is globally asymptotically stable. If
$\mathscr{R}_{0}\leq 1$, the disease-free steady state is globally asymptotically stable. If  $\mathscr{R}_{0}\gt 1$, the uniform persistence of phage-free model, as well as the existence of the phage-free steady state, are established. In a spatially homogeneous case, when
$\mathscr{R}_{0}\gt 1$, the uniform persistence of phage-free model, as well as the existence of the phage-free steady state, are established. In a spatially homogeneous case, when  $\ \;\widetilde{\!\!\!\mathscr{R}}_{0}\gt 1$, the global asymptotic stability of phage-free steady state and the uniform persistence of the phage-present model are discussed under some additional conditions. The mathematical approach here has wide applications in degenerate Partial Differential Equations.
$\ \;\widetilde{\!\!\!\mathscr{R}}_{0}\gt 1$, the global asymptotic stability of phage-free steady state and the uniform persistence of the phage-present model are discussed under some additional conditions. The mathematical approach here has wide applications in degenerate Partial Differential Equations.




Existence-uniqueness theory and small-data decay for a reaction-diffusion model of wildfire spread
- Part of
-
- Journal:
- Canadian Mathematical Bulletin / Volume 68 / Issue 4 / December 2025
- Published online by Cambridge University Press:
- 08 July 2025, pp. 1347-1358
- Print publication:
- December 2025
-
- Article
- Export citation
-
I examine some analytical properties of a nonlinear reaction-diffusion system that has been used to model the propagation of a wildfire. I establish global-in-time existence and uniqueness of bounded mild solutions to the Cauchy problem for this system given bounded initial data. In particular, this shows that the model does not allow for thermal blow-up. If the initial temperature and fuel density also satisfy certain integrability conditions, the spatial
 $L^2$-norms of these global solutions are uniformly bounded in time. Additionally, I use a bootstrap argument to show that small initial temperatures give rise to solutions that decay to zero as time goes to infinity, proving the existence of initial states that do not develop into traveling combustion waves.
$L^2$-norms of these global solutions are uniformly bounded in time. Additionally, I use a bootstrap argument to show that small initial temperatures give rise to solutions that decay to zero as time goes to infinity, proving the existence of initial states that do not develop into traveling combustion waves.

A free boundary problem of competition-diffusion system with Dirichlet boundary condition
- Part of
-
- Journal:
- Canadian Mathematical Bulletin , First View
- Published online by Cambridge University Press:
- 07 July 2025, pp. 1-20
-
- Article
- Export citation
-
In this article, we investigate a free boundary problem for the Lotka–Volterra model consisting of an invasive species with density u and a native species with density v in one dimension. We assume that v undergoes diffusion and growth in
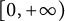 $[0,+\infty )$, and u invades into the environment with spreading front
$[0,+\infty )$, and u invades into the environment with spreading front  $x=h(t)$ satisfying free boundary condition
$x=h(t)$ satisfying free boundary condition  $h'(t)=-u_x(t,h(t))-\alpha $ for some decay rate
$h'(t)=-u_x(t,h(t))-\alpha $ for some decay rate 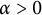 $\alpha>0$, this is caused by the bad environment at the boundary. When u is an inferior competitor,
$\alpha>0$, this is caused by the bad environment at the boundary. When u is an inferior competitor,  $u(t,x)$ and
$u(t,x)$ and 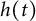 $h(t)$ tend to 0 within a finite time, while another specie
$h(t)$ tend to 0 within a finite time, while another specie 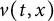 $v(t,x)$ tends to a stationary
$v(t,x)$ tends to a stationary 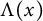 $\Lambda (x)$ defined on the half-line. When u is a superior competitor, we have a trichotomy result: spreading of u and vanishing of v (i.e., as
$\Lambda (x)$ defined on the half-line. When u is a superior competitor, we have a trichotomy result: spreading of u and vanishing of v (i.e., as 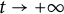 $t \to +\infty $,
$t \to +\infty $,  $h(t)$ goes to
$h(t)$ goes to  $+\infty $ and
$+\infty $ and 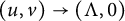 $(u,v)\to (\Lambda ,0)$); the transition case (i.e., as
$(u,v)\to (\Lambda ,0)$); the transition case (i.e., as 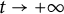 $t \to +\infty $,
$t \to +\infty $,  $(u,v)\to (w_\alpha , \eta _\alpha )$,
$(u,v)\to (w_\alpha , \eta _\alpha )$,  $h(t)$ tends to a finite point); vanishing of u and spreading of v (i.e.,
$h(t)$ tends to a finite point); vanishing of u and spreading of v (i.e., 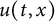 $u(t,x)$ and
$u(t,x)$ and  $h(t)$ tends to 0 within a finite time,
$h(t)$ tends to 0 within a finite time, 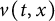 $v(t,x)$ converges to
$v(t,x)$ converges to 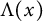 $\Lambda (x)$). Additionally, we show that this trichotomy result depends on the initial data
$\Lambda (x)$). Additionally, we show that this trichotomy result depends on the initial data  $u(0,x)$.
$u(0,x)$.
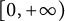


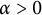

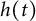
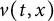
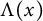
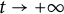


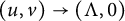
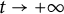


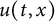

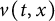
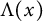

The evolution problem for the 1D nonlocal Fisher-KPP equation with a top hat kernel. Part 2. The initial-boundary value problem on a finite domain
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 04 July 2025, pp. 1-31
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In the second part of this series of papers, we address the same evolution problem that was considered in part 1 (see [16]), namely the nonlocal Fisher-KPP equation in one spatial dimension,
where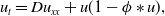 \begin{equation*} u_t = D u_{xx} + u(1-\phi *u), \end{equation*}
\begin{equation*} u_t = D u_{xx} + u(1-\phi *u), \end{equation*}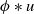 $\phi *u$ is a spatial convolution with the top hat kernel,
$\phi *u$ is a spatial convolution with the top hat kernel, 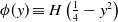 $\phi (y) \equiv H\left (\frac {1}{4}-y^2\right )$, except that now we modify this to an associated initial-boundary value problem on the finite spatial interval
$\phi (y) \equiv H\left (\frac {1}{4}-y^2\right )$, except that now we modify this to an associated initial-boundary value problem on the finite spatial interval 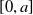 $[0,a]$ rather than the whole real line. Boundary conditions are required at the end points of the interval, and we address the situations when these are of either Dirichlet or Neumann type. This model is a natural extension of the classical Fisher-KPP model, with the introduction of the simplest possible nonlocal effect into the saturation term. Nonlocal reaction-diffusion models arise naturally in a variety of (frequently biological or ecological) contexts, and as such it is of fundamental interest to examine their properties in detail, and to compare and contrast these with the well known properties of the classical Fisher-KPP model.
$[0,a]$ rather than the whole real line. Boundary conditions are required at the end points of the interval, and we address the situations when these are of either Dirichlet or Neumann type. This model is a natural extension of the classical Fisher-KPP model, with the introduction of the simplest possible nonlocal effect into the saturation term. Nonlocal reaction-diffusion models arise naturally in a variety of (frequently biological or ecological) contexts, and as such it is of fundamental interest to examine their properties in detail, and to compare and contrast these with the well known properties of the classical Fisher-KPP model.
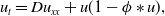
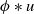
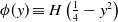
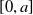
DELAYED HOPF BIFURCATIONS IN REACTION–DIFFUSION SYSTEMS IN TWO SPACE DIMENSIONS
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 67 / 2025
- Published online by Cambridge University Press:
- 13 May 2025, e19
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Pattern formation with jump discontinuity in a predator–prey model with Holling-II functional response
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 11 April 2025, pp. 1160-1182
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Spatio-temporal dynamics of a lattice prey–predator system with non-local diffusion in a periodic habitat
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 31 March 2025, pp. 1-43
-
- Article
- Export citation
Impact of prey-taxis on a harvested intraguild predation predator–prey model
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 10 March 2025, pp. 1183-1220
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
A critical non-homogeneous heat equation with weighted source
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 06 March 2025, pp. 1148-1159
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Some qualitative properties of radially symmetric solutions to the non-homogeneous heat equation with critical density and weighted source
are obtained, in the range of exponents \begin{align*} |x|^{-2}\partial _tu=\Delta u+|x|^{\sigma }u^p, \quad (x,t)\in {\mathbb {R}}^N\times (0,T), \end{align*}
\begin{align*} |x|^{-2}\partial _tu=\Delta u+|x|^{\sigma }u^p, \quad (x,t)\in {\mathbb {R}}^N\times (0,T), \end{align*} $p\gt 1$,
$p\gt 1$,  $\sigma \ge -2$. More precisely, we establish conditions fulfilled by the initial data in order for the solutions to either blow-up in finite time or decay to zero as
$\sigma \ge -2$. More precisely, we establish conditions fulfilled by the initial data in order for the solutions to either blow-up in finite time or decay to zero as  $t\to \infty$ and, in the latter case, we also deduce decay rates and large time behavior. In the limiting case
$t\to \infty$ and, in the latter case, we also deduce decay rates and large time behavior. In the limiting case  $\sigma =-2$, we prove the existence of non-trivial, non-negative solutions, in stark contrast to the homogeneous case. A transformation to a generalized Fisher–KPP equation is derived and employed in order to deduce these properties.
$\sigma =-2$, we prove the existence of non-trivial, non-negative solutions, in stark contrast to the homogeneous case. A transformation to a generalized Fisher–KPP equation is derived and employed in order to deduce these properties.





Global well-posedness and Turing–Hopf bifurcation of prey-taxis systems with hunting cooperation
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 24 February 2025, pp. 1121-1147
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Analysis of a model describing bacterial colony expansion in radial geometry driven by chemotaxis
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 5 / October 2025
- Published online by Cambridge University Press:
- 02 December 2024, pp. 910-947
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Global well-posedness and uniform boundedness of 2D urban crime models with nonlinear advection enhancement
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 5 / October 2025
- Published online by Cambridge University Press:
- 20 November 2024, pp. 897-909
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Global existence of a weak solution for a reaction–diffusion system in a porous medium with membrane conditions and mass control
- Part of
-
- Journal:
- Canadian Journal of Mathematics / Volume 77 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 20 November 2024, pp. 2006-2028
- Print publication:
- December 2025
-
- Article
- Export citation
-
In this paper, we prove the global exstence of weak solutions for a porous medium dynamics of m species moving between two domains separated by a zero-thickness membrane. On this membrane, Kedem–Katchalsky conditions are considered, and the study is characterized by natural structural conditions applied to the nonlinear reactive terms. The global existence is established under the assumption that these reactive terms are bounded in
 $L^1$. This problem has already been analyzed in the linear diffusion case by Ciavolella and Perthame in Ciavolella and Perthame (2021, Journal of Evolution Equations 21, 1513–1540). The present work constitutes an extension for nonlinear diffusion, particularly of the porous medium type, in the form
$L^1$. This problem has already been analyzed in the linear diffusion case by Ciavolella and Perthame in Ciavolella and Perthame (2021, Journal of Evolution Equations 21, 1513–1540). The present work constitutes an extension for nonlinear diffusion, particularly of the porous medium type, in the form  $\partial _t v_i - \Delta v_i^{r_i} = R_i$, for an exponent
$\partial _t v_i - \Delta v_i^{r_i} = R_i$, for an exponent  $r_i < 2$. The case
$r_i < 2$. The case  $r_i \geq 2$ remains an open problem. This paper is an adaptation of the ideas from Ciavolella and Perthame (2021, Journal of Evolution Equations 21, 1513–1540), with new strategies to overcome the appearance of nonlinearity and degeneracy in the diffusion term.
$r_i \geq 2$ remains an open problem. This paper is an adaptation of the ideas from Ciavolella and Perthame (2021, Journal of Evolution Equations 21, 1513–1540), with new strategies to overcome the appearance of nonlinearity and degeneracy in the diffusion term.




Fractional order for the transmission dynamics of coffee berry diseases (CBD)
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 20 November 2024, pp. 879-896
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
The evolution problem for the 1D nonlocal Fisher-KPP equation with a top hat kernel. Part 1. The Cauchy problem on the real line
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 25 October 2024, pp. 775-810
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We study the Cauchy problem on the real line for the nonlocal Fisher-KPP equation in one spatial dimension,
where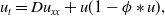 \begin{equation*} u_t = D u_{xx} + u(1-\phi *u), \end{equation*}
\begin{equation*} u_t = D u_{xx} + u(1-\phi *u), \end{equation*}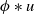 $\phi *u$ is a spatial convolution with the top hat kernel,
$\phi *u$ is a spatial convolution with the top hat kernel, 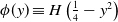 $\phi (y) \equiv H\left (\frac{1}{4}-y^2\right )$. After observing that the problem is globally well-posed, we demonstrate that positive, spatially periodic solutions bifurcate from the spatially uniform steady state solution
$\phi (y) \equiv H\left (\frac{1}{4}-y^2\right )$. After observing that the problem is globally well-posed, we demonstrate that positive, spatially periodic solutions bifurcate from the spatially uniform steady state solution 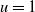 $u=1$ as the diffusivity,
$u=1$ as the diffusivity, 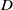 $D$, decreases through
$D$, decreases through 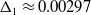 $\Delta _1 \approx 0.00297$ (the exact value is determined in Section 3). We explicitly construct these spatially periodic solutions as uniformly valid asymptotic approximations for
$\Delta _1 \approx 0.00297$ (the exact value is determined in Section 3). We explicitly construct these spatially periodic solutions as uniformly valid asymptotic approximations for 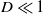 $D \ll 1$, over one wavelength, via the method of matched asymptotic expansions. These consist, at leading order, of regularly spaced, compactly supported regions with width of
$D \ll 1$, over one wavelength, via the method of matched asymptotic expansions. These consist, at leading order, of regularly spaced, compactly supported regions with width of 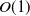 $O(1)$ where
$O(1)$ where 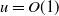 $u=O(1)$, separated by regions where
$u=O(1)$, separated by regions where  $u$ is exponentially small at leading order as
$u$ is exponentially small at leading order as 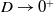 $D \to 0^+$. From numerical solutions, we find that for
$D \to 0^+$. From numerical solutions, we find that for 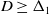 $D \geq \Delta _1$, permanent form travelling waves, with minimum wavespeed,
$D \geq \Delta _1$, permanent form travelling waves, with minimum wavespeed, 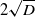 $2 \sqrt{D}$, are generated, whilst for
$2 \sqrt{D}$, are generated, whilst for 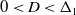 $0 \lt D \lt \Delta _1$, the wavefronts generated separate the regions where
$0 \lt D \lt \Delta _1$, the wavefronts generated separate the regions where 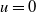 $u=0$ from a region where a steady periodic solution is created via a distinct periodic shedding mechanism acting immediately to the rear of the advancing front, with this mechanism becoming more pronounced with decreasing
$u=0$ from a region where a steady periodic solution is created via a distinct periodic shedding mechanism acting immediately to the rear of the advancing front, with this mechanism becoming more pronounced with decreasing 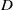 $D$. The structure of these transitional travelling wave forms is examined in some detail.
$D$. The structure of these transitional travelling wave forms is examined in some detail.
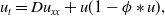
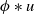
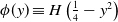
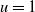
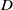
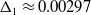
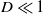
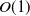
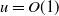

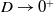
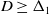
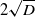
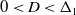
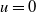
WELL-POSEDNESS RESULTS FOR GENERAL REACTION–DIFFUSION TRANSPORT OF OXYGEN IN ENCAPSULATED CELLS
- Part of
-
- Journal:
- Bulletin of the Australian Mathematical Society / Volume 111 / Issue 1 / February 2025
- Published online by Cambridge University Press:
- 18 October 2024, pp. 154-163
- Print publication:
- February 2025
-
- Article
- Export citation
Travelling wavefronts for the Belousov–Zhabotinsky system with non-local delayed interaction
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 10 October 2024, pp. 738-774
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Stabilization in a chemotaxis system modelling T-cell dynamics with simultaneous production and consumption of signals
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 3 / June 2025
- Published online by Cambridge University Press:
- 18 September 2024, pp. 570-583
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In a smoothly bounded domain
 $\Omega \subset \mathbb{R}^n$,
$\Omega \subset \mathbb{R}^n$,  $n\ge 1$, this manuscript considers the homogeneous Neumann boundary problem for the chemotaxis systemwith parameter
$n\ge 1$, this manuscript considers the homogeneous Neumann boundary problem for the chemotaxis systemwith parameter \begin{eqnarray*} \left \{ \begin{array}{l} u_t = \Delta u - \nabla \cdot (u\nabla v), \\[5pt] v_t = \Delta v + u - \alpha uv, \end{array} \right . \end{eqnarray*}
\begin{eqnarray*} \left \{ \begin{array}{l} u_t = \Delta u - \nabla \cdot (u\nabla v), \\[5pt] v_t = \Delta v + u - \alpha uv, \end{array} \right . \end{eqnarray*} $\alpha \gt 0$ and with coincident production and uptake of attractants, as recently emphasized by Dallaston et al. as relevant for the understanding of T-cell dynamics.
$\alpha \gt 0$ and with coincident production and uptake of attractants, as recently emphasized by Dallaston et al. as relevant for the understanding of T-cell dynamics.It is shown that there exists
 $\delta _\star =\delta _\star (n)\gt 0$ such that for any given
$\delta _\star =\delta _\star (n)\gt 0$ such that for any given  $\alpha \ge \frac{1}{\delta _\star }$ and for any suitably regular initial data satisfying
$\alpha \ge \frac{1}{\delta _\star }$ and for any suitably regular initial data satisfying  $v(\cdot, 0)\le \delta _\star$, this problem admits a unique classical solution that stabilizes to the constant equilibrium
$v(\cdot, 0)\le \delta _\star$, this problem admits a unique classical solution that stabilizes to the constant equilibrium  $(\frac{1}{|\Omega |}\int _\Omega u(\cdot, 0), \, \frac{1}{\alpha })$ in the large time limit.
$(\frac{1}{|\Omega |}\int _\Omega u(\cdot, 0), \, \frac{1}{\alpha })$ in the large time limit.








