Refine search
Actions for selected content:
384 results
Functional limit theorems for branching processes in a nearly degenerate varying environment
- Part of
-
- Journal:
- Journal of Applied Probability , First View
- Published online by Cambridge University Press:
- 20 May 2026, pp. 1-20
-
- Article
- Export citation
-
We investigate branching processes in a nearly degenerate varying environment, where the offspring distribution converges to the degenerate distribution at 1. Such processes die out almost surely; therefore, we either condition on non-extinction or add inhomogeneous immigration. Extending our one-dimensional limit results from Kevei and Kubatovics (2024), we derive functional limit theorems. In the former case, the limit process is a time-changed simple birth-and-death process on
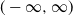 $(-\infty, \infty)$ conditioned on survival at 0, while in the latter, it is a time-changed stationary continuous-time Markov branching process with immigration.
$(-\infty, \infty)$ conditioned on survival at 0, while in the latter, it is a time-changed stationary continuous-time Markov branching process with immigration.
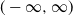
On McKean–Vlasov branching diffusion processes
- Part of
-
- Journal:
- Journal of Applied Probability , First View
- Published online by Cambridge University Press:
- 15 May 2026, pp. 1-25
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Remarks on Kolmogorov’s constant for simple branching processes
- Part of
-
- Journal:
- Journal of Applied Probability , First View
- Published online by Cambridge University Press:
- 28 April 2026, pp. 1-13
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Recent investigations have argued that there is a simple explicit representation for the Kolmogorov constant c associated with the subcritical Galton–Watson branching process. We exhibit examples showing that although this representation can be valid, it more often is not. Our work is presented in terms of the limiting conditional mean population size
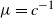 $\mu=c^{-1}$. The analogous quantity for the Markov branching process is denoted by
$\mu=c^{-1}$. The analogous quantity for the Markov branching process is denoted by 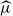 $\widehat\mu$. We show that the simple representation put forward for
$\widehat\mu$. We show that the simple representation put forward for 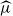 $\widehat\mu$ in fact is an upper bound that is attained only if the offspring-number probability-generating function is quadratic. The conditional mean
$\widehat\mu$ in fact is an upper bound that is attained only if the offspring-number probability-generating function is quadratic. The conditional mean 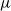 $\mu$ is the limit of a computable increasing sequence
$\mu$ is the limit of a computable increasing sequence 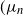 $(\mu_n$). Estimates of n are determined ensuring that, for any small positive number
$(\mu_n$). Estimates of n are determined ensuring that, for any small positive number  $\varepsilon$,
$\varepsilon$, 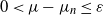 $0\lt\mu-\mu_n\le \varepsilon$.
$0\lt\mu-\mu_n\le \varepsilon$.
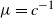
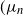

A star is born: Explosive Crump–Mode–Jagers branching processes
- Part of
-
- Journal:
- Journal of Applied Probability , First View
- Published online by Cambridge University Press:
- 13 April 2026, pp. 1-19
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Strong survival and extinction for branching random walks via new order for generating functions
- Part of
-
- Journal:
- Journal of Applied Probability , First View
- Published online by Cambridge University Press:
- 05 March 2026, pp. 1-19
-
- Article
- Export citation
-
We consider general discrete-time multitype branching processes on a countable set X. According to these processes, a particle of type
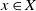 $x\in X$ generates a random number of children and chooses their type in X, not necessarily independently nor with the same law for different parent types. We introduce a new type of stochastic ordering of multitype branching processes, generalising the germ order introduced by Hutchcroft, which relies on the generating function of the process. We prove that given two multitype branching processes with laws
$x\in X$ generates a random number of children and chooses their type in X, not necessarily independently nor with the same law for different parent types. We introduce a new type of stochastic ordering of multitype branching processes, generalising the germ order introduced by Hutchcroft, which relies on the generating function of the process. We prove that given two multitype branching processes with laws 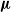 ${\boldsymbol{\mu}}$ and
${\boldsymbol{\mu}}$ and  ${\boldsymbol{\nu}}$ respectively, with
${\boldsymbol{\nu}}$ respectively, with 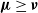 ${\boldsymbol{\mu}}\ge{\boldsymbol{\nu}}$, then in every set where there is survival according to
${\boldsymbol{\mu}}\ge{\boldsymbol{\nu}}$, then in every set where there is survival according to  ${\boldsymbol{\nu}}$, there is also survival according to
${\boldsymbol{\nu}}$, there is also survival according to 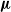 ${\boldsymbol{\mu}}$. Moreover, in every set where there is strong survival according to
${\boldsymbol{\mu}}$. Moreover, in every set where there is strong survival according to  ${\boldsymbol{\nu}}$, there is also strong survival according to
${\boldsymbol{\nu}}$, there is also strong survival according to 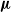 ${\boldsymbol{\mu}}$, provided that the supremum of the global extinction probabilities for the
${\boldsymbol{\mu}}$, provided that the supremum of the global extinction probabilities for the  ${\boldsymbol{\nu}}$ process, taken over all starting points x, is strictly smaller than 1. New conditions for survival and strong survival for inhomogeneous multitype branching processes are provided. We also extend a result of Moyal which claims that, under some conditions, the global extinction probability for a multitype branching process is the only fixed point of its generating function, whose supremum over all starting coordinates may be smaller than 1.
${\boldsymbol{\nu}}$ process, taken over all starting points x, is strictly smaller than 1. New conditions for survival and strong survival for inhomogeneous multitype branching processes are provided. We also extend a result of Moyal which claims that, under some conditions, the global extinction probability for a multitype branching process is the only fixed point of its generating function, whose supremum over all starting coordinates may be smaller than 1.
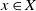
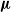

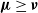

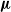


The largest subcritical component in inhomogeneous random graphs of preferential attachment type
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 35 / Issue 3 / May 2026
- Published online by Cambridge University Press:
- 04 March 2026, pp. 414-432
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
De Finetti’s control for refracted skew Brownian motion
- Part of
-
- Journal:
- Advances in Applied Probability , First View
- Published online by Cambridge University Press:
- 03 March 2026, pp. 1-42
-
- Article
- Export citation
On asymptotic behavior of time of extinction of critical bisexual branching process in random environment
- Part of
-
- Journal:
- Journal of Applied Probability , First View
- Published online by Cambridge University Press:
- 02 March 2026, pp. 1-13
-
- Article
- Export citation
-
We consider a critical bisexual branching process in a random environment generated by independent and identically distributed random variables. Assuming that the process starts with a large number of pairs N, we prove that its extinction time is of order
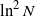 $\ln^2 N$. Interestingly, this result is valid for a general class of mating functions. Among these are the functions describing the monogamous and polygamous behavior of couples, as well as the function reducing the bisexual branching process to the simple one.
$\ln^2 N$. Interestingly, this result is valid for a general class of mating functions. Among these are the functions describing the monogamous and polygamous behavior of couples, as well as the function reducing the bisexual branching process to the simple one.
Multitype branching processes with immigration generated by point processes
- Part of
-
- Journal:
- Journal of Applied Probability , First View
- Published online by Cambridge University Press:
- 12 January 2026, pp. 1-21
-
- Article
- Export citation
Branching particle systems with mutually catalytic interactions
- Part of
-
- Journal:
- Advances in Applied Probability , First View
- Published online by Cambridge University Press:
- 11 December 2025, pp. 1-38
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We study a continuous-time mutually catalytic branching model on the
 $\mathbb{Z}^{d}$. The model describes the behavior of two different populations of particles, performing random walk on the lattice in the presence of branching, that is, each particle dies at a certain rate and is replaced by a random number of offspring. The branching rate of a particle in one population is proportional to the number of particles of another population at the same site. We study the long time behavior for this model, in particular, coexistence and noncoexistence of two populations in the long run. Finally, we construct a sequence of renormalized processes and use duality techniques to investigate its limiting behavior.
$\mathbb{Z}^{d}$. The model describes the behavior of two different populations of particles, performing random walk on the lattice in the presence of branching, that is, each particle dies at a certain rate and is replaced by a random number of offspring. The branching rate of a particle in one population is proportional to the number of particles of another population at the same site. We study the long time behavior for this model, in particular, coexistence and noncoexistence of two populations in the long run. Finally, we construct a sequence of renormalized processes and use duality techniques to investigate its limiting behavior.

Mixed-state branching evolution for cell division models
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 63 / Issue 1 / March 2026
- Published online by Cambridge University Press:
- 29 September 2025, pp. 135-156
- Print publication:
- March 2026
-
- Article
- Export citation
-
We prove a scaling limit theorem for two-type Galton–Watson branching processes with interaction. The limit theorem gives rise to a class of mixed-state branching processes with interaction used to simulate evolution for cell division affected by parasites. Such processes can also be obtained by the pathwise-unique solution to a stochastic equation system. Moreover, we present sufficient conditions for extinction with probability 1 and the exponential ergodicity in the
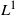 $L^1$-Wasserstein distance of such processes in some cases.
$L^1$-Wasserstein distance of such processes in some cases.
Two-type branching processes with immigration, and the structured coalescents
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 63 / Issue 1 / March 2026
- Published online by Cambridge University Press:
- 29 September 2025, pp. 32-60
- Print publication:
- March 2026
-
- Article
- Export citation
Conditioning Bienaymé–Galton–Watson trees to have large sub-populations
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 58 / Issue 1 / March 2026
- Published online by Cambridge University Press:
- 10 September 2025, pp. 345-386
- Print publication:
- March 2026
-
- Article
- Export citation
Quasi-stationary distributions for subcritical branching Markov chains
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 58 / Issue 1 / March 2026
- Published online by Cambridge University Press:
- 10 September 2025, pp. 425-471
- Print publication:
- March 2026
-
- Article
- Export citation
-
Consider a subcritical branching Markov chain. Let
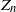 $Z_n$ denote the counting measure of particles of generation n. Under some conditions, we give a probabilistic proof for the existence of the Yaglom limit of
$Z_n$ denote the counting measure of particles of generation n. Under some conditions, we give a probabilistic proof for the existence of the Yaglom limit of  $(Z_n)_{n\in\mathbb{N}}$ by the moment method, based on the spinal decomposition and the many-to-few formula. As a result, we give explicit integral representations of all quasi-stationary distributions of
$(Z_n)_{n\in\mathbb{N}}$ by the moment method, based on the spinal decomposition and the many-to-few formula. As a result, we give explicit integral representations of all quasi-stationary distributions of  $(Z_n)_{n\in\mathbb{N}}$, whose proofs are direct and probabilistic, and do not rely on Martin boundary theory.
$(Z_n)_{n\in\mathbb{N}}$, whose proofs are direct and probabilistic, and do not rely on Martin boundary theory.
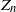


The distance on the slightly supercritical random series–parallel graph
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 58 / Issue 1 / March 2026
- Published online by Cambridge University Press:
- 09 September 2025, pp. 80-121
- Print publication:
- March 2026
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider the random series–parallel graph introduced by Hambly and Jordan (2004 Adv. Appl. Probab. 36, 824–838), which is a hierarchical graph with a parameter
 $p\in [0, \, 1]$. The graph is built recursively: at each step, every edge in the graph is either replaced with probability p by a series of two edges, or with probability
$p\in [0, \, 1]$. The graph is built recursively: at each step, every edge in the graph is either replaced with probability p by a series of two edges, or with probability 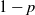 $1-p$ by two parallel edges, and the replacements are independent of each other and of everything up to then. At the nth step of the recursive procedure, the distance between the extremal points on the graph is denoted by
$1-p$ by two parallel edges, and the replacements are independent of each other and of everything up to then. At the nth step of the recursive procedure, the distance between the extremal points on the graph is denoted by  $D_n (p)$. It is known that
$D_n (p)$. It is known that  $D_n(p)$ possesses a phase transition at
$D_n(p)$ possesses a phase transition at  $p=p_c \;:\!=\;\frac{1}{2}$; more precisely,
$p=p_c \;:\!=\;\frac{1}{2}$; more precisely,  $\frac{1}{n}\log {{\mathbb{E}}}[D_n(p)] \to \alpha(p)$ when
$\frac{1}{n}\log {{\mathbb{E}}}[D_n(p)] \to \alpha(p)$ when  $n \to \infty$, with
$n \to \infty$, with  $\alpha(p) >0$ for
$\alpha(p) >0$ for  $p>p_c$ and
$p>p_c$ and  $\alpha(p)=0$ for
$\alpha(p)=0$ for  $p\le p_c$. We study the exponent
$p\le p_c$. We study the exponent  $\alpha(p)$ in the slightly supercritical regime
$\alpha(p)$ in the slightly supercritical regime 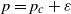 $p=p_c+\varepsilon$. Our main result says that as
$p=p_c+\varepsilon$. Our main result says that as  $\varepsilon\to 0^+$,
$\varepsilon\to 0^+$,  $\alpha(p_c+\varepsilon)$ behaves like
$\alpha(p_c+\varepsilon)$ behaves like 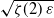 $\sqrt{\zeta(2) \, \varepsilon}$, where
$\sqrt{\zeta(2) \, \varepsilon}$, where  $\zeta(2) \;:\!=\; \frac{\pi^2}{6}$.
$\zeta(2) \;:\!=\; \frac{\pi^2}{6}$.

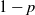










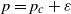


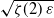

Negative moments of the CREM partition function in the high-temperature regime
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 4 / December 2025
- Published online by Cambridge University Press:
- 29 July 2025, pp. 1623-1641
- Print publication:
- December 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The continuous random energy model (CREM) was introduced by Bovier and Kurkova in 2004 as a toy model of disordered systems. Among other things, their work indicates that there exists a critical point
 $\beta_\mathrm{c}$ such that the partition function exhibits a phase transition. The present work focuses on the high-temperature regime where
$\beta_\mathrm{c}$ such that the partition function exhibits a phase transition. The present work focuses on the high-temperature regime where  $\beta<\beta_\mathrm{c}$. We show that, for all
$\beta<\beta_\mathrm{c}$. We show that, for all  $\beta<\beta_\mathrm{c}$ and for all
$\beta<\beta_\mathrm{c}$ and for all  $s>0$, the negative s moment of the CREM partition function is comparable with the expectation of the CREM partition function to the power of
$s>0$, the negative s moment of the CREM partition function is comparable with the expectation of the CREM partition function to the power of  $-s$, up to constants that are independent of N.
$-s$, up to constants that are independent of N.





The replicator coalescent
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 4 / December 2025
- Published online by Cambridge University Press:
- 24 June 2025, pp. 1534-1548
- Print publication:
- December 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider a stochastic model, called the replicator coalescent, describing a system of blocks of k different types that undergo pairwise mergers at rates depending on the block types: with rate
 $C_{ij}\geq 0$ blocks of type i and j merge, resulting in a single block of type i. The replicator coalescent can be seen as a generalisation of Kingman’s coalescent death chain in a multi-type setting, although without an underpinning exchangeable partition structure. The name is derived from a remarkable connection between the instantaneous dynamics of this multi-type coalescent when issued from an arbitrarily large number of blocks, and the so-called replicator equations from evolutionary game theory. By dilating time arbitrarily close to zero, we see that initially, on coming down from infinity, the replicator coalescent behaves like the solution to a certain replicator equation. Thereafter, stochastic effects are felt and the process evolves more in the spirit of a multi-type death chain.
$C_{ij}\geq 0$ blocks of type i and j merge, resulting in a single block of type i. The replicator coalescent can be seen as a generalisation of Kingman’s coalescent death chain in a multi-type setting, although without an underpinning exchangeable partition structure. The name is derived from a remarkable connection between the instantaneous dynamics of this multi-type coalescent when issued from an arbitrarily large number of blocks, and the so-called replicator equations from evolutionary game theory. By dilating time arbitrarily close to zero, we see that initially, on coming down from infinity, the replicator coalescent behaves like the solution to a certain replicator equation. Thereafter, stochastic effects are felt and the process evolves more in the spirit of a multi-type death chain.

Ergodic theorem for branching markov chains indexed by trees with arbitrary shape
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 3 / September 2025
- Published online by Cambridge University Press:
- 09 June 2025, pp. 1089-1104
- Print publication:
- September 2025
-
- Article
- Export citation
-
We prove an ergodic theorem for Markov chains indexed by the Ulam–Harris–Neveu tree over large subsets with arbitrary shape under two assumptions: (i) with high probability, two vertices in the large subset are far from each other, and (ii) with high probability, those two vertices have their common ancestor close to the root. The assumption on the common ancestor can be replaced by some regularity assumption on the Markov transition kernel. We verify that these assumptions are satisfied for some usual trees. Finally, with Markov chain Monte Carlo considerations in mind, we prove that when the underlying Markov chain is stationary and reversible, the Markov chain, that is the line graph, yields minimal variance for the empirical average estimator among trees with a given number of nodes. In doing so, we prove that the Hosoya–Wiener polynomial is minimized over
 $[{-}1,1]$ by the line graph among trees of a given size.
$[{-}1,1]$ by the line graph among trees of a given size.

Epidemic models with digital and manual contact tracing
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 4 / December 2025
- Published online by Cambridge University Press:
- 09 June 2025, pp. 1430-1455
- Print publication:
- December 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We analyse a Markovian SIR epidemic model where individuals either recover naturally or are diagnosed, leading to isolation and potential contact tracing. Our focus is on digital contact tracing via a tracing app, considering both its standalone use and its combination with manual tracing. We prove that as the population size n grows large, the epidemic process converges to a limiting process, which, unlike with typical epidemic models, is not a branching process due to dependencies created by contact tracing. However, by grouping to-be-traced individuals into macro-individuals, we derive a multi-type branching process interpretation, allowing computation of the reproduction number R. This is then converted to an individual reproduction number
 $R^\mathrm{(ind)}$, which, in contrast to R, decays monotonically with the fraction of app-users, while both share the same threshold at 1. Finally, we compare digital (only) contact tracing and manual (only) contact tracing, proving that the critical fraction of app-users,
$R^\mathrm{(ind)}$, which, in contrast to R, decays monotonically with the fraction of app-users, while both share the same threshold at 1. Finally, we compare digital (only) contact tracing and manual (only) contact tracing, proving that the critical fraction of app-users,  $\pi_{\mathrm{c}}$, required for
$\pi_{\mathrm{c}}$, required for  $R=1$ is higher than the critical fraction manually contact-traced,
$R=1$ is higher than the critical fraction manually contact-traced,  $p_{\mathrm{c}}$, for manual tracing.
$p_{\mathrm{c}}$, for manual tracing.




Asymptotic behaviors of subcritical branching killed Brownian motion with drift
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 57 / Issue 4 / December 2025
- Published online by Cambridge University Press:
- 26 May 2025, pp. 1484-1509
- Print publication:
- December 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In this paper, we study asymptotic behaviors of a subcritical branching Brownian motion with drift
 $-\rho$, killed upon exiting
$-\rho$, killed upon exiting  $(0, \infty)$, and offspring distribution
$(0, \infty)$, and offspring distribution  $\{p_k{:}\; k\ge 0\}$. Let
$\{p_k{:}\; k\ge 0\}$. Let  $\widetilde{\zeta}^{-\rho}$ be the extinction time of this subcritical branching killed Brownian motion,
$\widetilde{\zeta}^{-\rho}$ be the extinction time of this subcritical branching killed Brownian motion,  $\widetilde{M}_t^{-\rho}$ the maximal position of all the particles alive at time t and
$\widetilde{M}_t^{-\rho}$ the maximal position of all the particles alive at time t and  $\widetilde{M}^{-\rho}:\!=\max_{t\ge 0}\widetilde{M}_t^{-\rho}$ the all-time maximal position. Let
$\widetilde{M}^{-\rho}:\!=\max_{t\ge 0}\widetilde{M}_t^{-\rho}$ the all-time maximal position. Let  $\mathbb{P}_x$ be the law of this subcritical branching killed Brownian motion when the initial particle is located at
$\mathbb{P}_x$ be the law of this subcritical branching killed Brownian motion when the initial particle is located at  $x\in (0,\infty)$. Under the assumption
$x\in (0,\infty)$. Under the assumption  $\sum_{k=1}^\infty k ({\log}\; k) p_k <\infty$, we establish the decay rates of
$\sum_{k=1}^\infty k ({\log}\; k) p_k <\infty$, we establish the decay rates of  $\mathbb{P}_x(\widetilde{\zeta}^{-\rho}>t)$ and
$\mathbb{P}_x(\widetilde{\zeta}^{-\rho}>t)$ and  $\mathbb{P}_x(\widetilde{M}^{-\rho}>y)$ as t and y respectively tend to
$\mathbb{P}_x(\widetilde{M}^{-\rho}>y)$ as t and y respectively tend to  $\infty$. We also establish the decay rate of
$\infty$. We also establish the decay rate of  $\mathbb{P}_x(\widetilde{M}_t^{-\rho}> z(t,\rho))$ as
$\mathbb{P}_x(\widetilde{M}_t^{-\rho}> z(t,\rho))$ as  $t\to\infty$, where
$t\to\infty$, where  $z(t,\rho)=\sqrt{t}z-\rho t$ for
$z(t,\rho)=\sqrt{t}z-\rho t$ for  $\rho\leq 0$ and
$\rho\leq 0$ and  $z(t,\rho)=z$ for
$z(t,\rho)=z$ for  $\rho>0$. As a consequence, we obtain a Yaglom-type limit theorem.
$\rho>0$. As a consequence, we obtain a Yaglom-type limit theorem.


















