Refine search
Actions for selected content:
238141 results in Physics and Astronomy
Superhydrophobic surfaces with recirculating interfacial flow due to surfactants are ‘effectively’ immobilized
-
- Journal:
- Journal of Fluid Mechanics / Volume 956 / 10 February 2023
- Published online by Cambridge University Press:
- 30 January 2023, R3
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Predicting radial profiles for jets with arbitrary buoyancy
-
- Journal:
- Journal of Fluid Mechanics / Volume 956 / 10 February 2023
- Published online by Cambridge University Press:
- 30 January 2023, A9
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Drop impact onto a substrate wetted by another liquid: corona detachment from the wall film
-
- Journal:
- Journal of Fluid Mechanics / Volume 956 / 10 February 2023
- Published online by Cambridge University Press:
- 30 January 2023, A10
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
FLM volume 955 Cover and Front matter
-
- Journal:
- Journal of Fluid Mechanics / Volume 955 / 25 January 2023
- Published online by Cambridge University Press:
- 30 January 2023, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Regime transitions in stratified shear flows: the link between horizontal and inclined ducts
-
- Journal:
- Journal of Fluid Mechanics / Volume 956 / 10 February 2023
- Published online by Cambridge University Press:
- 26 January 2023, A4
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We present the analytical solution for the two-dimensional velocity and density fields within an approximation for laminar stratified inclined duct (SID) flows where diffusion dominates over inertia in the along-channel momentum equation but is negligible in the density transport equation. We refer to this approximation as the hydrostatic/gravitational/viscous in momentum and advective in density (HGV-A) approximation due to the leading balances in the governing equations. The analytical solution is valid for laminar flows in a two-layer configuration in the limit of long ducts. The non-dimensional volume flux within the HGV-A approximation is given by
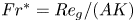 $Fr^* ={{Re}}_g/(AK)$, which is a control parameter with
$Fr^* ={{Re}}_g/(AK)$, which is a control parameter with 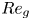 ${{Re}}_g$ the gravitational Reynolds number,
${{Re}}_g$ the gravitational Reynolds number, 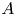 $A$ the aspect ratio of the duct and
$A$ the aspect ratio of the duct and 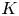 $K$ a geometrical parameter that depends on the tilt of the duct and is obtained from the analytical solution. This analytical solution was validated against results from laboratory experiments, and allows us to gain new insight into the dynamics and properties of SID flows. Most importantly, constant values of
$K$ a geometrical parameter that depends on the tilt of the duct and is obtained from the analytical solution. This analytical solution was validated against results from laboratory experiments, and allows us to gain new insight into the dynamics and properties of SID flows. Most importantly, constant values of 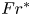 $Fr^*$ describe, in both horizontal and inclined ducts, the transitions between increasingly turbulent flow regimes: from laminar flow, to interfacial waves, to intermittent turbulence and sustained turbulence.
$Fr^*$ describe, in both horizontal and inclined ducts, the transitions between increasingly turbulent flow regimes: from laminar flow, to interfacial waves, to intermittent turbulence and sustained turbulence.
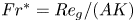
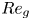
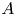
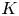
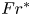
Chapter 10 - Epilepsy as a Complex Network Disorder
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 135-152
-
- Chapter
- Export citation
Chapter 3 - Transcriptomic and Epigenomic Approaches for Epilepsy
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 19-40
-
- Chapter
- Export citation
Chapter 9 - A Neuroimaging Network-Level Approach to Drug-Resistant Epilepsy
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 127-134
-
- Chapter
- Export citation
Chapter 1 - Introduction
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 1-4
-
- Chapter
- Export citation
The unsteady aerodynamics of insect wings with rotational stroke accelerations, a systematic numerical study – ERRATUM
-
- Journal:
- Journal of Fluid Mechanics / Volume 956 / 10 February 2023
- Published online by Cambridge University Press:
- 26 January 2023, E1
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Index
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 153-160
-
- Chapter
- Export citation
Chapter 2 - Systems Biology Approaches to the Genetic Complexity of Epilepsy
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 5-18
-
- Chapter
- Export citation
Chapter 6 - The Baseline and Epileptiform EEG
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 72-85
-
- Chapter
- Export citation
Chapter 4 - Phenomenological Mesoscopic Models for Seizure Activity
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 41-60
-
- Chapter
- Export citation
Contents
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp v-v
-
- Chapter
- Export citation

Acoustic resonance mechanism for axisymmetric screech modes of underexpanded jets impinging on an inclined plate
-
- Journal:
- Journal of Fluid Mechanics / Volume 956 / 10 February 2023
- Published online by Cambridge University Press:
- 26 January 2023, A2
-
- Article
- Export citation
-
In this paper, the acoustic resonance mechanism for different axisymmetric screech modes of the underexpanded jets that impinge on an inclined plate is investigated experimentally. The ideally expanded Mach number of jets (
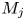 $M_j$) ranges from 1.05 to 1.56. The nozzle-to-plate distance at the jet axis and the impingement angle are respectively set as 5.0
$M_j$) ranges from 1.05 to 1.56. The nozzle-to-plate distance at the jet axis and the impingement angle are respectively set as 5.0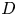 $D$ and
$D$ and 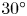 $30^{\circ }$, where
$30^{\circ }$, where 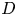 $D$ is the nozzle exit diameter. The acoustic results show that the
$D$ is the nozzle exit diameter. The acoustic results show that the 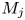 $M_j$ range for the A2 screech mode of impinging jets is broader than that of underexpanded free jets, and a new axisymmetric screech mode A3 appears. With the increase of
$M_j$ range for the A2 screech mode of impinging jets is broader than that of underexpanded free jets, and a new axisymmetric screech mode A3 appears. With the increase of 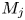 $M_j$, the effect of the impinging plate on the shock cell structures of jets becomes obvious gradually, and the second suboptimal peaks are evident in the axial wavenumber spectra of mean shock structures. The coherent flow structures at screech frequencies are extracted from time-resolved schlieren images via the spectral proper orthogonal decomposition (SPOD). The axial wavenumber spectra of the selected SPOD modes suggest that the A1, A2 and A3 screech modes are respectively closed by the guided jet modes that are energized by the interactions between the Kelvin–Helmholtz wavepacket and the first three shock wavenumber peaks. The upstream- and downstream-propagating waves that constitute the screech feedback loop are analysed by applying wavenumber filters to the wavenumber spectra of SPOD modes. The frequencies of these three screech modes can be predicted by the phase constraints between the nozzle exit and the rear edge of the third shock cell. For the A3 mode, the inclined plate invades the third shock cell with the increase of
$M_j$, the effect of the impinging plate on the shock cell structures of jets becomes obvious gradually, and the second suboptimal peaks are evident in the axial wavenumber spectra of mean shock structures. The coherent flow structures at screech frequencies are extracted from time-resolved schlieren images via the spectral proper orthogonal decomposition (SPOD). The axial wavenumber spectra of the selected SPOD modes suggest that the A1, A2 and A3 screech modes are respectively closed by the guided jet modes that are energized by the interactions between the Kelvin–Helmholtz wavepacket and the first three shock wavenumber peaks. The upstream- and downstream-propagating waves that constitute the screech feedback loop are analysed by applying wavenumber filters to the wavenumber spectra of SPOD modes. The frequencies of these three screech modes can be predicted by the phase constraints between the nozzle exit and the rear edge of the third shock cell. For the A3 mode, the inclined plate invades the third shock cell with the increase of 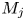 $M_j$, and the phase constraint cannot be satisfied at the lower side of the jets, which leads the A3 mode to fade away. The present results suggest that external boundaries can modulate the frequency and mode of jet screech by changing the axial spacings of shock cells.
$M_j$, and the phase constraint cannot be satisfied at the lower side of the jets, which leads the A3 mode to fade away. The present results suggest that external boundaries can modulate the frequency and mode of jet screech by changing the axial spacings of shock cells.
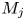
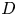
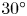
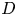
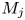
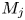
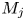
Copyright page
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp iv-iv
-
- Chapter
- Export citation
Contributors
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp vi-viii
-
- Chapter
- Export citation
Chapter 7 - Neuronal Approaches to Epilepsy
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 86-98
-
- Chapter
- Export citation
Chapter 5 - Personalized Network Modeling in Epilepsy
-
-
- Book:
- A Complex Systems Approach to Epilepsy
- Published online:
- 06 January 2023
- Print publication:
- 26 January 2023, pp 61-71
-
- Chapter
- Export citation
