Refine listing
Actions for selected content:
1723 results in 60Jxx
Second-order differential equations with random perturbations and small parameters
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 147 / Issue 4 / August 2017
- Published online by Cambridge University Press:
- 05 July 2017, pp. 763-779
- Print publication:
- August 2017
-
- Article
- Export citation
Subcritical Sevastyanov branching processes with nonhomogeneous Poisson immigration
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 22 June 2017, pp. 569-587
- Print publication:
- June 2017
-
- Article
- Export citation
On the range of the transient frog model on ℤ
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 49 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 26 June 2017, pp. 327-343
- Print publication:
- June 2017
-
- Article
- Export citation
Coupling any number of balls in the infinite-bin model
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 22 June 2017, pp. 540-549
- Print publication:
- June 2017
-
- Article
- Export citation
Multitype infinite-allele branching processes in continuous time
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 22 June 2017, pp. 550-568
- Print publication:
- June 2017
-
- Article
- Export citation
On some mixing times for nonreversible finite Markov chains
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 22 June 2017, pp. 627-637
- Print publication:
- June 2017
-
- Article
- Export citation
On the large-time behaviour of the solution of a stochastic differential equation driven by a Poisson point process
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 49 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 26 June 2017, pp. 344-367
- Print publication:
- June 2017
-
- Article
- Export citation
Optimal bulking threshold of batch service queues
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 22 June 2017, pp. 409-423
- Print publication:
- June 2017
-
- Article
- Export citation
Path to survival for the critical branching processes in a random environment
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 22 June 2017, pp. 588-602
- Print publication:
- June 2017
-
- Article
- Export citation
Nash equilibrium in nonzero-sum games of optimal stopping for Brownian motion
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 49 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 26 June 2017, pp. 430-445
- Print publication:
- June 2017
-
- Article
- Export citation
Limit theorems for some branching measure-valued processes
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 49 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 26 June 2017, pp. 549-580
- Print publication:
- June 2017
-
- Article
-
- You have access
- Export citation
Ergodicity of Markov chain Monte Carlo with reversible proposal
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 22 June 2017, pp. 638-654
- Print publication:
- June 2017
-
- Article
- Export citation
Marshall–Olkin distributions, subordinators, efficient simulation, and applications to credit risk
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 49 / Issue 2 / June 2017
- Published online by Cambridge University Press:
- 26 June 2017, pp. 481-514
- Print publication:
- June 2017
-
- Article
- Export citation
ON BOUNDED-TYPE THIN LOCAL SETS OF THE TWO-DIMENSIONAL GAUSSIAN FREE FIELD
- Part of
-
- Journal:
- Journal of the Institute of Mathematics of Jussieu / Volume 18 / Issue 3 / May 2019
- Published online by Cambridge University Press:
- 27 April 2017, pp. 591-618
- Print publication:
- May 2019
-
- Article
- Export citation
-
We study certain classes of local sets of the two-dimensional Gaussian free field (GFF) in a simply connected domain, and their relation to the conformal loop ensemble
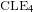 $\text{CLE}_{4}$ and its variants. More specifically, we consider bounded-type thin local sets (BTLS), where thin means that the local set is small in size, and bounded type means that the harmonic function describing the mean value of the field away from the local set is bounded by some deterministic constant. We show that a local set is a BTLS if and only if it is contained in some nested version of the
$\text{CLE}_{4}$ and its variants. More specifically, we consider bounded-type thin local sets (BTLS), where thin means that the local set is small in size, and bounded type means that the harmonic function describing the mean value of the field away from the local set is bounded by some deterministic constant. We show that a local set is a BTLS if and only if it is contained in some nested version of the 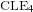 $\text{CLE}_{4}$ carpet, and prove that all BTLS are necessarily connected to the boundary of the domain. We also construct all possible BTLS for which the corresponding harmonic function takes only two prescribed values and show that all these sets (and this includes the case of
$\text{CLE}_{4}$ carpet, and prove that all BTLS are necessarily connected to the boundary of the domain. We also construct all possible BTLS for which the corresponding harmonic function takes only two prescribed values and show that all these sets (and this includes the case of 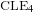 $\text{CLE}_{4}$) are in fact measurable functions of the GFF.
$\text{CLE}_{4}$) are in fact measurable functions of the GFF.
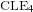
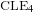
Surprise Probabilities in Markov Chains
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 26 / Issue 4 / July 2017
- Published online by Cambridge University Press:
- 16 March 2017, pp. 603-627
-
- Article
- Export citation
-
In a Markov chain started at a state x, the hitting time τ(y) is the first time that the chain reaches another state y. We study the probability
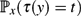 $\mathbb{P}_x(\tau(y) = t)$ that the first visit to y occurs precisely at a given time t. Informally speaking, the event that a new state is visited at a large time t may be considered a ‘surprise’. We prove the following three bounds.
$\mathbb{P}_x(\tau(y) = t)$ that the first visit to y occurs precisely at a given time t. Informally speaking, the event that a new state is visited at a large time t may be considered a ‘surprise’. We prove the following three bounds.-
• In any Markov chain with n states,
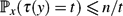 $\mathbb{P}_x(\tau(y) = t) \le {n}/{t}$ .
$\mathbb{P}_x(\tau(y) = t) \le {n}/{t}$ . -
• In a reversible chain with n states,
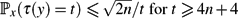 $\mathbb{P}_x(\tau(y) = t) \le {\sqrt{2n}}/{t}$ for $t \ge 4n + 4$ .
$\mathbb{P}_x(\tau(y) = t) \le {\sqrt{2n}}/{t}$ for $t \ge 4n + 4$ . -
• For random walk on a simple graph with n ≥ 2 vertices,
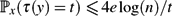 $\mathbb{P}_x(\tau(y) = t) \le 4e \log(n)/t$ .
$\mathbb{P}_x(\tau(y) = t) \le 4e \log(n)/t$ .
To prove the bound for random walk on graphs, we establish the following estimate conjectured by Aldous, Ding and Oveis-Gharan (private communication): for random walk on an n-vertex graph, for every initial vertex x,
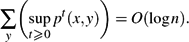 $$\sum_y \biggl( \sup_{t \ge 0} p^t(x, y) \biggr) = O(\log n). $$
$$\sum_y \biggl( \sup_{t \ge 0} p^t(x, y) \biggr) = O(\log n). $$ -
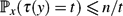
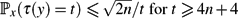
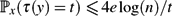
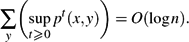
ON THE CONVERGENCE OF DISCRETE PROCESSES WITH MULTIPLE INDEPENDENT VARIABLES
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 58 / Issue 3-4 / April 2017
- Published online by Cambridge University Press:
- 06 March 2017, pp. 379-385
-
- Article
-
- You have access
- Export citation
Local limits of Galton–Watson trees conditioned on the number of protected nodes
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 1 / March 2017
- Published online by Cambridge University Press:
- 04 April 2017, pp. 55-65
- Print publication:
- March 2017
-
- Article
- Export citation
On the central management of risk networks
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 49 / Issue 1 / March 2017
- Published online by Cambridge University Press:
- 17 March 2017, pp. 221-237
- Print publication:
- March 2017
-
- Article
-
- You have access
- Export citation
Statistical inference for partially observed branching processes with immigration
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 54 / Issue 1 / March 2017
- Published online by Cambridge University Press:
- 04 April 2017, pp. 82-95
- Print publication:
- March 2017
-
- Article
- Export citation
Extreme events of Markov chains
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 49 / Issue 1 / March 2017
- Published online by Cambridge University Press:
- 17 March 2017, pp. 134-161
- Print publication:
- March 2017
-
- Article
- Export citation
