Refine search
Actions for selected content:
26063 results in Abstract analysis
Propagation of anisotropic Gabor wave front sets
- Part of
- Miscellaneous topics - Partial differential equations
- Partial differential equations
- Distributions, generalized functions, distribution spaces
- Groups and semigroups of linear operators, their generalizations and applications
- Partial differential equations on manifolds; differential operators
- General quantum mechanics and problems of quantization
- Integral, integro-differential, and pseudodifferential operators
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 67 / Issue 3 / August 2024
- Published online by Cambridge University Press:
- 22 April 2024, pp. 674-698
-
- Article
- Export citation
Behaviour of the minimum degree throughout the
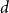 ${\textit{d}}$-process
${\textit{d}}$-process
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 33 / Issue 5 / September 2024
- Published online by Cambridge University Press:
- 22 April 2024, pp. 564-582
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The
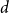 $d$-process generates a graph at random by starting with an empty graph with
$d$-process generates a graph at random by starting with an empty graph with  $n$ vertices, then adding edges one at a time uniformly at random among all pairs of vertices which have degrees at most
$n$ vertices, then adding edges one at a time uniformly at random among all pairs of vertices which have degrees at most 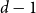 $d-1$ and are not mutually joined. We show that, in the evolution of a random graph with
$d-1$ and are not mutually joined. We show that, in the evolution of a random graph with  $n$ vertices under the
$n$ vertices under the 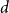 $d$-process with
$d$-process with 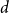 $d$ fixed, with high probability, for each
$d$ fixed, with high probability, for each 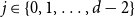 $j \in \{0,1,\dots,d-2\}$, the minimum degree jumps from
$j \in \{0,1,\dots,d-2\}$, the minimum degree jumps from 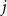 $j$ to
$j$ to 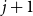 $j+1$ when the number of steps left is on the order of
$j+1$ when the number of steps left is on the order of 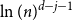 $\ln (n)^{d-j-1}$. This answers a question of Ruciński and Wormald. More specifically, we show that, when the last vertex of degree
$\ln (n)^{d-j-1}$. This answers a question of Ruciński and Wormald. More specifically, we show that, when the last vertex of degree 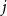 $j$ disappears, the number of steps left divided by
$j$ disappears, the number of steps left divided by 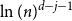 $\ln (n)^{d-j-1}$ converges in distribution to the exponential random variable of mean
$\ln (n)^{d-j-1}$ converges in distribution to the exponential random variable of mean 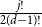 $\frac{j!}{2(d-1)!}$; furthermore, these
$\frac{j!}{2(d-1)!}$; furthermore, these 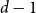 $d-1$ distributions are independent.
$d-1$ distributions are independent.

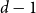

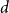
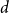
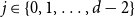
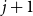
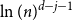
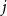
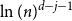
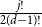
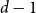
Uniform propagation of chaos for a dollar exchange econophysics model
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 1 / February 2025
- Published online by Cambridge University Press:
- 22 April 2024, pp. 27-39
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Simplicial volume of manifolds with amenable fundamental group at infinity
- Part of
-
- Journal:
- Glasgow Mathematical Journal / Volume 66 / Issue 3 / September 2024
- Published online by Cambridge University Press:
- 22 April 2024, pp. 563-570
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We show that for
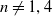 $n \neq 1,4$, the simplicial volume of an inward tame triangulable open
$n \neq 1,4$, the simplicial volume of an inward tame triangulable open  $n$-manifold
$n$-manifold  $M$ with amenable fundamental group at infinity at each end is finite; moreover, we show that if also
$M$ with amenable fundamental group at infinity at each end is finite; moreover, we show that if also 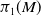 $\pi _1(M)$ is amenable, then the simplicial volume of
$\pi _1(M)$ is amenable, then the simplicial volume of 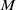 $M$ vanishes. We show that the same result holds for finitely-many-ended triangulable manifolds which are simply connected at infinity.
$M$ vanishes. We show that the same result holds for finitely-many-ended triangulable manifolds which are simply connected at infinity.

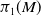
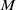
Singer conjecture for varieties with semismall Albanese map and residually finite fundamental group
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 156 / Issue 1 / February 2026
- Published online by Cambridge University Press:
- 19 April 2024, pp. 122-128
- Print publication:
- February 2026
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Macroscopic limit of a Fokker-Planck model of swarming rigid bodies
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 2 / April 2025
- Published online by Cambridge University Press:
- 19 April 2024, pp. 349-397
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider self-propelled rigid bodies interacting through local body-attitude alignment modelled by stochastic differential equations. We derive a hydrodynamic model of this system at large spatio-temporal scales and particle numbers in any dimension
 $n \geq 3$. This goal was already achieved in dimension
$n \geq 3$. This goal was already achieved in dimension  $n=3$ or in any dimension
$n=3$ or in any dimension  $n \geq 3$ for a different system involving jump processes. However, the present work corresponds to huge conceptual and technical gaps compared with earlier ones. The key difficulty is to determine an auxiliary but essential object, the generalised collision invariant. We achieve this aim by using the geometrical structure of the rotation group, namely its maximal torus, Cartan subalgebra and Weyl group as well as other concepts of representation theory and Weyl’s integration formula. The resulting hydrodynamic model appears as a hyperbolic system whose coefficients depend on the generalised collision invariant.
$n \geq 3$ for a different system involving jump processes. However, the present work corresponds to huge conceptual and technical gaps compared with earlier ones. The key difficulty is to determine an auxiliary but essential object, the generalised collision invariant. We achieve this aim by using the geometrical structure of the rotation group, namely its maximal torus, Cartan subalgebra and Weyl group as well as other concepts of representation theory and Weyl’s integration formula. The resulting hydrodynamic model appears as a hyperbolic system whose coefficients depend on the generalised collision invariant.



Pushforward of structure sheaf and virtual global generation
- Part of
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 67 / Issue 3 / August 2024
- Published online by Cambridge University Press:
- 18 April 2024, pp. 663-673
-
- Article
- Export citation
-
Let
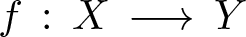 $f\,:\,X\,\longrightarrow \,Y$ be a generically smooth morphism between irreducible smooth projective curves over an algebraically closed field of arbitrary characteristic. We prove that the vector bundle
$f\,:\,X\,\longrightarrow \,Y$ be a generically smooth morphism between irreducible smooth projective curves over an algebraically closed field of arbitrary characteristic. We prove that the vector bundle 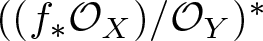 $((f_*{\mathcal O}_X)/{\mathcal O}_Y)^*$ is virtually globally generated. Moreover,
$((f_*{\mathcal O}_X)/{\mathcal O}_Y)^*$ is virtually globally generated. Moreover, 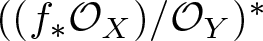 $((f_*{\mathcal O}_X)/{\mathcal O}_Y)^*$ is ample if and only if f is genuinely ramified.
$((f_*{\mathcal O}_X)/{\mathcal O}_Y)^*$ is ample if and only if f is genuinely ramified.
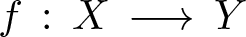
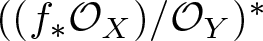
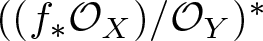
ON THE DISTINCTION OF IWAHORI-SPHERICAL DISCRETE SERIES REPRESENTATIONS
- Part of
-
- Journal:
- Journal of the Institute of Mathematics of Jussieu / Volume 24 / Issue 1 / January 2025
- Published online by Cambridge University Press:
- 17 April 2024, pp. 1-15
- Print publication:
- January 2025
-
- Article
- Export citation
-
Let
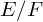 $E/F$ be a quadratic unramified extension of non-archimedean local fields and
$E/F$ be a quadratic unramified extension of non-archimedean local fields and  $\mathbb H$ a simply connected semisimple algebraic group defined and split over F. We establish general results (multiplicities, test vectors) on
$\mathbb H$ a simply connected semisimple algebraic group defined and split over F. We establish general results (multiplicities, test vectors) on 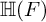 ${\mathbb H} (F)$-distinguished Iwahori-spherical representations of
${\mathbb H} (F)$-distinguished Iwahori-spherical representations of 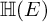 ${\mathbb H} (E)$. For discrete series Iwahori-spherical representations of
${\mathbb H} (E)$. For discrete series Iwahori-spherical representations of 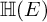 ${\mathbb H} (E)$, we prove a numerical criterion of
${\mathbb H} (E)$, we prove a numerical criterion of 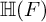 ${\mathbb H} (F)$-distinction. As an application, we classify the
${\mathbb H} (F)$-distinction. As an application, we classify the 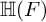 ${\mathbb H} (F)$-distinguished discrete series representations of
${\mathbb H} (F)$-distinguished discrete series representations of 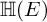 ${\mathbb H} (E)$ corresponding to degree
${\mathbb H} (E)$ corresponding to degree 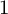 $1$ characters of the Iwahori-Hecke algebra.
$1$ characters of the Iwahori-Hecke algebra.
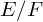

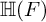
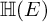
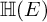
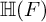
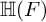
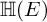
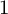
The homotopy decomposition of the suspension of a non-simply-connected five-manifold
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 156 / Issue 1 / February 2026
- Published online by Cambridge University Press:
- 17 April 2024, pp. 93-121
- Print publication:
- February 2026
-
- Article
- Export citation
-
In this paper we determine the homotopy types of the reduced suspension space of certain connected orientable closed smooth $five$
 -manifolds. As applications, we compute the reduced $K$
-manifolds. As applications, we compute the reduced $K$ -groups of $M$
-groups of $M$ and show that the suspension map between the third cohomotopy set $\pi ^3(M)$
and show that the suspension map between the third cohomotopy set $\pi ^3(M)$ and the fourth cohomotopy set $\pi ^4(\Sigma M)$
and the fourth cohomotopy set $\pi ^4(\Sigma M)$ is a bijection.
is a bijection.





Maximal subgroups of a family of iterated monodromy groups
- Part of
-
- Journal:
- Glasgow Mathematical Journal / Volume 66 / Issue 3 / September 2024
- Published online by Cambridge University Press:
- 17 April 2024, pp. 541-562
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The Basilica group is a well-known 2-generated weakly branch, but not branch, group acting on the binary rooted tree. Recently, a more general form of the Basilica group has been investigated by Petschick and Rajeev, which is an
 $s$-generated weakly branch, but not branch, group that acts on the
$s$-generated weakly branch, but not branch, group that acts on the  $m$-adic tree, for
$m$-adic tree, for 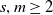 $s,m\ge 2$. A larger family of groups, which contains these generalised Basilica groups, is the family of iterated monodromy groups. With the new developments by Francoeur, the study of the existence of maximal subgroups of infinite index has been extended from branch groups to weakly branch groups. Here we show that a subfamily of iterated monodromy groups, which more closely resemble the generalised Basilica groups, have maximal subgroups only of finite index.
$s,m\ge 2$. A larger family of groups, which contains these generalised Basilica groups, is the family of iterated monodromy groups. With the new developments by Francoeur, the study of the existence of maximal subgroups of infinite index has been extended from branch groups to weakly branch groups. Here we show that a subfamily of iterated monodromy groups, which more closely resemble the generalised Basilica groups, have maximal subgroups only of finite index.


KOBAYASHI-OCHIAI’S FINITENESS THEOREM FOR ORBIFOLD PAIRS OF GENERAL TYPE
- Part of
-
- Journal:
- Journal of the Institute of Mathematics of Jussieu / Volume 23 / Issue 6 / November 2024
- Published online by Cambridge University Press:
- 17 April 2024, pp. 2713-2732
- Print publication:
- November 2024
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
A generalization of Bondy’s pancyclicity theorem
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 33 / Issue 5 / September 2024
- Published online by Cambridge University Press:
- 17 April 2024, pp. 554-563
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The bipartite independence number of a graph
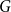 $G$, denoted as
$G$, denoted as 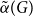 $\tilde \alpha (G)$, is the minimal number
$\tilde \alpha (G)$, is the minimal number 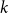 $k$ such that there exist positive integers
$k$ such that there exist positive integers  $a$ and
$a$ and 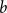 $b$ with
$b$ with 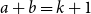 $a+b=k+1$ with the property that for any two disjoint sets
$a+b=k+1$ with the property that for any two disjoint sets 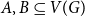 $A,B\subseteq V(G)$ with
$A,B\subseteq V(G)$ with 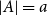 $|A|=a$ and
$|A|=a$ and 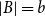 $|B|=b$, there is an edge between
$|B|=b$, there is an edge between 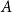 $A$ and
$A$ and 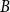 $B$. McDiarmid and Yolov showed that if
$B$. McDiarmid and Yolov showed that if 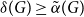 $\delta (G)\geq \tilde \alpha (G)$ then
$\delta (G)\geq \tilde \alpha (G)$ then 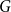 $G$ is Hamiltonian, extending the famous theorem of Dirac which states that if
$G$ is Hamiltonian, extending the famous theorem of Dirac which states that if 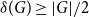 $\delta (G)\geq |G|/2$ then
$\delta (G)\geq |G|/2$ then 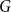 $G$ is Hamiltonian. In 1973, Bondy showed that, unless
$G$ is Hamiltonian. In 1973, Bondy showed that, unless 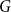 $G$ is a complete bipartite graph, Dirac’s Hamiltonicity condition also implies pancyclicity, i.e., existence of cycles of all the lengths from
$G$ is a complete bipartite graph, Dirac’s Hamiltonicity condition also implies pancyclicity, i.e., existence of cycles of all the lengths from 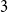 $3$ up to
$3$ up to  $n$. In this paper, we show that
$n$. In this paper, we show that 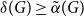 $\delta (G)\geq \tilde \alpha (G)$ implies that
$\delta (G)\geq \tilde \alpha (G)$ implies that 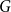 $G$ is pancyclic or that
$G$ is pancyclic or that 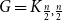 $G=K_{\frac{n}{2},\frac{n}{2}}$, thus extending the result of McDiarmid and Yolov, and generalizing the classic theorem of Bondy.
$G=K_{\frac{n}{2},\frac{n}{2}}$, thus extending the result of McDiarmid and Yolov, and generalizing the classic theorem of Bondy.

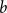
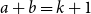
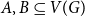
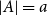
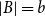
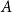
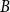
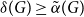
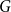
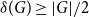
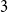

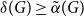
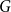
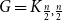
Slow passage through the Busse balloon – predicting steps on the Eckhaus staircase
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 1 / February 2025
- Published online by Cambridge University Press:
- 16 April 2024, pp. 1-26
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Sharp conditions for the validity of the Bourgain–Brezis–Mironescu formula
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 156 / Issue 1 / February 2026
- Published online by Cambridge University Press:
- 16 April 2024, pp. 69-92
- Print publication:
- February 2026
-
- Article
- Export citation
-
Following the seminal paper by Bourgain, Brezis, and Mironescu, we focus on the asymptotic behaviour of some nonlocal functionals that, for each $u\in L^2(\mathbb {R}^N)$
 , are defined as the double integrals of weighted, squared difference quotients of $u$
, are defined as the double integrals of weighted, squared difference quotients of $u$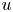 . Given a family of weights $\{\rho _{\varepsilon} \}$
. Given a family of weights $\{\rho _{\varepsilon} \}$ , $\varepsilon \in (0,\,1)$
, $\varepsilon \in (0,\,1)$ , we devise sufficient and necessary conditions on $\{\rho _{\varepsilon} \}$
, we devise sufficient and necessary conditions on $\{\rho _{\varepsilon} \}$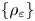 for the associated nonlocal functionals to converge as $\varepsilon \to 0$
for the associated nonlocal functionals to converge as $\varepsilon \to 0$ to a variant of the Dirichlet integral. Finally, some comparison between our result and the existing literature is provided.
to a variant of the Dirichlet integral. Finally, some comparison between our result and the existing literature is provided.




A finite-volume scheme for fractional diffusion on bounded domains
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 2 / April 2025
- Published online by Cambridge University Press:
- 16 April 2024, pp. 398-418
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Large Péclet number forced convection from a circular wire in a uniform stream: hybrid approximations at small Reynolds numbers
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 35 / Issue 6 / December 2024
- Published online by Cambridge University Press:
- 16 April 2024, pp. 823-834
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider heat or mass transport from a circular cylinder under a uniform crossflow at small Reynolds numbers,
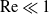 $\mathrm{Re}\ll 1$. This problem has been thwarted in the past by limitations inherent in the classical analyses of the singular flow problem, which have used asymptotic expansions in inverse powers of
$\mathrm{Re}\ll 1$. This problem has been thwarted in the past by limitations inherent in the classical analyses of the singular flow problem, which have used asymptotic expansions in inverse powers of 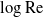 $\log \mathrm{Re}$. We here make use of the hybrid approximation of Kropinski, Ward & Keller [(1995) SIAM J. Appl. Math. 55, 1484], based upon a robust asymptotic expansion in powers of
$\log \mathrm{Re}$. We here make use of the hybrid approximation of Kropinski, Ward & Keller [(1995) SIAM J. Appl. Math. 55, 1484], based upon a robust asymptotic expansion in powers of 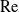 $\mathrm{Re}$. In that approximation, the “inner” streamfunction is provided by the product of a pre-factor
$\mathrm{Re}$. In that approximation, the “inner” streamfunction is provided by the product of a pre-factor 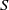 $S$, a slowly varying function of
$S$, a slowly varying function of 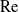 $\mathrm{Re}$, with a
$\mathrm{Re}$, with a 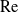 $\mathrm{Re}$-independent “canonical” solution of a simple mathematical form. The pre-factor, in turn, is determined as an implicit function of
$\mathrm{Re}$-independent “canonical” solution of a simple mathematical form. The pre-factor, in turn, is determined as an implicit function of 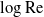 $\log \mathrm{Re}$ via asymptotic matching with a numerical solution of the nonlinear single-scaled “outer” problem, where the cylinder appears as a point singularity. We exploit the hybrid approximation to analyse the transport problem in the limit of large Péclet number,
$\log \mathrm{Re}$ via asymptotic matching with a numerical solution of the nonlinear single-scaled “outer” problem, where the cylinder appears as a point singularity. We exploit the hybrid approximation to analyse the transport problem in the limit of large Péclet number, 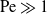 $\mathrm{Pe}\gg 1$. In that limit, transport is restricted to a narrow boundary layer about the cylinder surface – a province contained within the inner region of the flow problem. With
$\mathrm{Pe}\gg 1$. In that limit, transport is restricted to a narrow boundary layer about the cylinder surface – a province contained within the inner region of the flow problem. With 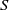 $S$ appearing as a parameter, a similarity solution is readily constructed for the boundary-layer problem. It provides the Nusselt number as
$S$ appearing as a parameter, a similarity solution is readily constructed for the boundary-layer problem. It provides the Nusselt number as 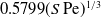 $0.5799(S\,\mathrm{Pe})^{1/3}$. This asymptotic prediction is in remarkably close agreement with that of the numerical solution of the exact problem [Dennis, Hudson & Smith (1968) Phys. Fluids 11, 933] even for moderate
$0.5799(S\,\mathrm{Pe})^{1/3}$. This asymptotic prediction is in remarkably close agreement with that of the numerical solution of the exact problem [Dennis, Hudson & Smith (1968) Phys. Fluids 11, 933] even for moderate 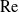 $\mathrm{Re}$-values.
$\mathrm{Re}$-values.
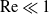
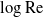
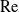
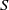
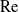
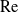
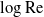
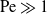
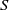
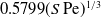
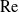
Linear actions of $\mathbb {Z}/p\times \mathbb {Z}/p$
 on $S^{2n-1}\times S^{2n-1}$
on $S^{2n-1}\times S^{2n-1}$
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 154 / Issue 6 / December 2024
- Published online by Cambridge University Press:
- 16 April 2024, pp. 1699-1712
- Print publication:
- December 2024
-
- Article
- Export citation
-
For an odd prime $p$
 , we consider free actions of $(\mathbb {Z}_{/{p}})^2$
, we consider free actions of $(\mathbb {Z}_{/{p}})^2$ on $S^{2n-1}\times S^{2n-1}$
on $S^{2n-1}\times S^{2n-1}$ given by linear actions of $(\mathbb {Z}_{/{p}})^2$
given by linear actions of $(\mathbb {Z}_{/{p}})^2$ on $\mathbb {R}^{4n}$
on $\mathbb {R}^{4n}$ . Simple examples include a lens space cross a lens space, but $k$
. Simple examples include a lens space cross a lens space, but $k$ -invariant calculations show that other quotients exist. Using the tools of Postnikov towers and surgery theory, the quotients are classified up to homotopy by the $k$
-invariant calculations show that other quotients exist. Using the tools of Postnikov towers and surgery theory, the quotients are classified up to homotopy by the $k$ -invariants and up to homeomorphism by the Pontrjagin classes. We will present these results and demonstrate how to calculate the $k$
-invariants and up to homeomorphism by the Pontrjagin classes. We will present these results and demonstrate how to calculate the $k$ -invariants and the Pontrjagin classes from the rotation numbers.
-invariants and the Pontrjagin classes from the rotation numbers.










A remark on a conjecture on the symmetric Gaussian problem
- Part of
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 67 / Issue 2 / May 2024
- Published online by Cambridge University Press:
- 15 April 2024, pp. 643-661
-
- Article
- Export citation
Twisted Blanchfield pairings and twisted signatures III: Applications
- Part of
-
- Journal:
- Glasgow Mathematical Journal / Volume 66 / Issue 3 / September 2024
- Published online by Cambridge University Press:
- 15 April 2024, pp. 501-540
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
This paper describes how to compute algorithmically certain twisted signature invariants of a knot
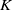 $K$ using twisted Blanchfield forms. An illustration of the algorithm is implemented on
$K$ using twisted Blanchfield forms. An illustration of the algorithm is implemented on 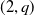 $(2,q)$-torus knots. Additionally, using satellite formulas for these invariants, we also show how to obstruct the sliceness of certain iterated torus knots.
$(2,q)$-torus knots. Additionally, using satellite formulas for these invariants, we also show how to obstruct the sliceness of certain iterated torus knots.
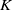
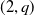
On the fractional powers of a Schrödinger operator with a Hardy-type potential
- Part of
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 67 / Issue 2 / May 2024
- Published online by Cambridge University Press:
- 12 April 2024, pp. 460-507
-
- Article
- Export citation
