Refine search
Actions for selected content:
238452 results in Physics and Astronomy

The Rise of Econophysics
- A Connected History of Two Disciplines
-
- Published online:
- 01 April 2026
- Print publication:
- 30 April 2026
-
- Element
- Export citation
Influence of Tsallis q-entropy on electron-impact ionisation in a non-extensive plasma
-
- Journal:
- Journal of Plasma Physics / Volume 92 / Issue 2 / April 2026
- Published online by Cambridge University Press:
- 01 April 2026, E37
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Experimental study of argon gas breakdown with symmetric and asymmetric electrode configurations
-
- Journal:
- Journal of Plasma Physics / Volume 92 / Issue 2 / April 2026
- Published online by Cambridge University Press:
- 01 April 2026, E44
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Paschen’s law relates the breakdown voltage (
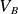 $V_B$) of a gas to the product of the gas pressure (
$V_B$) of a gas to the product of the gas pressure (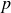 $p$) and the inter-electrode distance (
$p$) and the inter-electrode distance (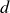 $d$), predicting a characteristic minimum breakdown voltage at a specific
$d$), predicting a characteristic minimum breakdown voltage at a specific 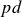 $pd$ value. In this study, the roles of electrode configurations (symmetric and asymmetric) and inter-electrode spacing on gas breakdown processes (or Paschen’s law) are examined. Numerous sets of experiments are performed with both symmetric and asymmetric electrode configurations of different sizes to obtain Paschen curves at different inter-electrode distances. The experimentally obtained Paschen’s curves for different electrode configurations are fitted using a proposed modified empirical relation for
$pd$ value. In this study, the roles of electrode configurations (symmetric and asymmetric) and inter-electrode spacing on gas breakdown processes (or Paschen’s law) are examined. Numerous sets of experiments are performed with both symmetric and asymmetric electrode configurations of different sizes to obtain Paschen curves at different inter-electrode distances. The experimentally obtained Paschen’s curves for different electrode configurations are fitted using a proposed modified empirical relation for 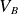 $V_B$, incorporating variable power-law dependencies and fitting parameters to better capture the observed deviations. Upon closer inspection, we observed that the breakdown voltage (
$V_B$, incorporating variable power-law dependencies and fitting parameters to better capture the observed deviations. Upon closer inspection, we observed that the breakdown voltage ( $V_B$) and the corresponding
$V_B$) and the corresponding 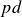 $pd$ value (
$pd$ value (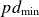 $pd_{\min}$) are influenced by both electrode configurations and inter-electrode discharge gap. The variation in
$pd_{\min}$) are influenced by both electrode configurations and inter-electrode discharge gap. The variation in 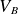 $V_B$ and
$V_B$ and 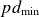 $pd_{\min}$ for different electrode configurations is explained by analysing the electric field distributions between the electrodes (cathode and anode) for an applied voltage.
$pd_{\min}$ for different electrode configurations is explained by analysing the electric field distributions between the electrodes (cathode and anode) for an applied voltage.
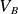
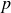
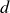
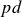
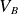

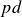
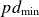
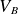
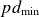

Channel confinement enhances group cohesion of self-propelled flapping plates via flow-mediated interactions
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 31 March 2026, A38
-
- Article
- Export citation
-
The hydrodynamic interactions involved in the self-organisation phenomenon in biological systems are not fully understood and have attracted significant attention. A previous study (Peng et al. 2018 J. Fluid Mech., vol. 853, pp. 587–600) found that, arranged in an unbounded fluid, the largest cluster of self-propelled bodies in tandem, capable of spontaneously forming an ordered configuration, consists of eight swimmers. Here, we numerically investigate the collective behaviour of multiple self-propelled plates in tandem within a channel of width
 $H$, confined by two parallel walls. These plates are driven by harmonic flapping motions of uniform frequency and amplitude. Results demonstrate for the first time that the channel confinement significantly enhances group cohesion, with up to 72 individuals self-organising into ordered configurations at an optimal channel width. We observe two stable configurations: a hybrid mode with subgroups (typically at smaller channel widths) and a regular mode with sparse configuration. In large regular-mode groups, the vortex fields downstream exhibit spatial periodicity, conforming to Rosenhead’s stability criterion for confined vortex streets (vortex spacing
$H$, confined by two parallel walls. These plates are driven by harmonic flapping motions of uniform frequency and amplitude. Results demonstrate for the first time that the channel confinement significantly enhances group cohesion, with up to 72 individuals self-organising into ordered configurations at an optimal channel width. We observe two stable configurations: a hybrid mode with subgroups (typically at smaller channel widths) and a regular mode with sparse configuration. In large regular-mode groups, the vortex fields downstream exhibit spatial periodicity, conforming to Rosenhead’s stability criterion for confined vortex streets (vortex spacing 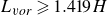 $L_{v\textit{or}} \geqslant 1.419H$). This theoretical alignment explains both the observed upper channel-width limit (
$L_{v\textit{or}} \geqslant 1.419H$). This theoretical alignment explains both the observed upper channel-width limit ( $H \approx 4.0{-}4.5$) for large-scale cohesion and the robust order in the regular mode. The plates may adopt spontaneously a ‘vortex-slalom’ path, reducing the drag force and energy consumption while maximising stability. Deviation from this path results in a spring-like restoring force, promptly returning the plate to equilibrium.
$H \approx 4.0{-}4.5$) for large-scale cohesion and the robust order in the regular mode. The plates may adopt spontaneously a ‘vortex-slalom’ path, reducing the drag force and energy consumption while maximising stability. Deviation from this path results in a spring-like restoring force, promptly returning the plate to equilibrium.

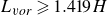

The impact of viscosity on the linear growth of the sausage and magneto-Rayleigh–Taylor instabilities in imploding cylindrical liners
-
- Journal:
- Journal of Plasma Physics / Volume 92 / Issue 2 / April 2026
- Published online by Cambridge University Press:
- 31 March 2026, E43
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Magnetised liner inertial fusion (MagLIF) has attracted attention in the past decade for its high obtained Lawson triple products and prospects to scale to ignition. In this work, we investigate the effect of viscosity on the sausage instability and magneto-Rayleigh–Taylor instability (MRTI) in conditions relevant for MagLIF implosions. First, we quantify the amount of damping that viscosity has on instability growth by deriving an expression for the ratio between viscous and inviscid growth rates. This expression is parameterised by a single non-dimensional number: the Galilei number
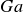 $Ga$, which measures the ratio of gravitational and viscous forces. We discuss in detail the physical intuition
$Ga$, which measures the ratio of gravitational and viscous forces. We discuss in detail the physical intuition 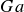 $Ga$ provides on instability growth. The derived growth rates are then validated against FLASH simulations. We then calculate a critical viscosity threshold
$Ga$ provides on instability growth. The derived growth rates are then validated against FLASH simulations. We then calculate a critical viscosity threshold  $\eta _{c}$ required for viscosity to dampen the instability growth rate by 5 %. From this analysis, we show that, for drive currents relevant to laboratory MagLIF experiments (of the order of tens of MA), this critical viscosity threshold is much greater than realistic liner viscosity values except for the shortest perturbation wavelength regimes. We conclude that viscosity does not play a significant role in the initial linear growth of the sausage instability and MRTI in MagLIF liners, but our results motivate future investigation into effects of viscosity in nonlinear and high temperature regimes.
$\eta _{c}$ required for viscosity to dampen the instability growth rate by 5 %. From this analysis, we show that, for drive currents relevant to laboratory MagLIF experiments (of the order of tens of MA), this critical viscosity threshold is much greater than realistic liner viscosity values except for the shortest perturbation wavelength regimes. We conclude that viscosity does not play a significant role in the initial linear growth of the sausage instability and MRTI in MagLIF liners, but our results motivate future investigation into effects of viscosity in nonlinear and high temperature regimes.


Propagation and interaction of gravity currents induced by successive release of two lock fluids
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 31 March 2026, A37
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
This study investigates the dynamics of sequentially released gravity currents in a lock-exchange configuration consisting of two lock fluids using high-resolution numerical simulations and compares them with the classical single-lock exchange. The results demonstrate that, in a two-lock-fluid configuration, the overtaking of the lighter lock current by the heavier lock current alters the current’s front dynamics, leading to complex velocity transitions. Before overtaking occurs, the front propagation is slower than in the classical single-lock-fluid case because of lower density contrast, but after overtaking, the heavier lock fluid accumulates at the head region of the current and enhances its speed. The head of the current is primarily dominated by streamwise velocity vectors, which directly influence the front propagation speed. The body of the current exhibits significant components of streamwise and wall-normal velocities, characterised by large eddies and Kelvin–Helmholtz billows at the interface, enhancing entrainment and mixing. In contrast, the tail region consists of small-scale eddies, which contribute to viscous dissipation, gradually reducing the current’s momentum. A parametric study of the two-lock-fluid configuration, conducted to investigate the effects of non-dimensionalised time of delay in the release of heavier lock fluid,
 $t_R^*$, the ratio of densities of ambient fluid to heavier lock fluid,
$t_R^*$, the ratio of densities of ambient fluid to heavier lock fluid,  $\gamma _2=\rho _a/\rho _2$, and the non-dimensionalised time of overtaking by the heavier lock fluid,
$\gamma _2=\rho _a/\rho _2$, and the non-dimensionalised time of overtaking by the heavier lock fluid,  $t_O^*$, revealed a nonlinear relationship. As local Reynolds number decreases
$t_O^*$, revealed a nonlinear relationship. As local Reynolds number decreases  $(Re_l\lesssim 10{\,}000)$, the relationship becomes nonlinear due to weaker buoyancy forcing of the heavier current travelling in the wake of the lighter current. However, for
$(Re_l\lesssim 10{\,}000)$, the relationship becomes nonlinear due to weaker buoyancy forcing of the heavier current travelling in the wake of the lighter current. However, for 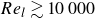 $Re_l\gtrsim 10{\,}000$, this relationship becomes linear.
$Re_l\gtrsim 10{\,}000$, this relationship becomes linear.





The influence of body anisotropy on wake characteristics and enstrophy production for prolate ellipsoids at ReD = 10 000
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 31 March 2026, A32
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
A least-action principle for visco-resistive Hall magnetohydrodynamics with metriplectic reformulation
-
- Journal:
- Journal of Plasma Physics / Volume 92 / Issue 2 / April 2026
- Published online by Cambridge University Press:
- 31 March 2026, E42
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Variability of mean-flow reversals in stably stratified fluids with stochastic wave forcing
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 31 March 2026, A29
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Lift force models for a spherical particle immersed in linear and quadratic shear flows
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 31 March 2026, A31
-
- Article
- Export citation
-
The lift force models for a particle in wall-bounded linear shear flow have been extensively investigated; however, the influence of the curvature of the velocity profile (
 ${\textit{Sg}}$) on the lift force at finite slip Reynolds numbers (
${\textit{Sg}}$) on the lift force at finite slip Reynolds numbers ( ${\textit{Re}}$) remains unexplored. In the present work, direct numerical simulations (DNS) are performed to investigate the lift on a spherical particle in unbounded linear shear flow, single-wall-bounded linear shear flow and Poiseuille flow. Based on our DNS data, we first extend the existing unbounded and single-wall-bounded linear shear-slip-induced lift models to higher non-dimensional shear rates (
${\textit{Re}}$) remains unexplored. In the present work, direct numerical simulations (DNS) are performed to investigate the lift on a spherical particle in unbounded linear shear flow, single-wall-bounded linear shear flow and Poiseuille flow. Based on our DNS data, we first extend the existing unbounded and single-wall-bounded linear shear-slip-induced lift models to higher non-dimensional shear rates ( $|Sr|=2.5$) for
$|Sr|=2.5$) for  $0.1\leqslant Re\leqslant 20$. Based on the empirical model for Couette flow or the analytical model for unbounded Poiseuille flow, the lift models are then modified to account for the curvature effect of the parabolic velocity profile, which reduce to the linear shear-slip-induced lift models in the high
$0.1\leqslant Re\leqslant 20$. Based on the empirical model for Couette flow or the analytical model for unbounded Poiseuille flow, the lift models are then modified to account for the curvature effect of the parabolic velocity profile, which reduce to the linear shear-slip-induced lift models in the high 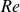 ${\textit{Re}}$ and low
${\textit{Re}}$ and low  ${\textit{Sg}}$ limits. We also modify the rotation-induced lift model of linear shear flow to account for the parabolic shear effect, which causes lift enhancement for the leading particle and lift attenuation for the lagging particle in the Poiseuille flow, compared to the linear shear case. This lift attenuation may give rise to an inverse Magnus force at low slip Reynolds numbers. In addition, the model for the particle free rotation rate for the Poiseuille flow is established by correcting the one for the linear shear flow, providing a more accurate prediction of the torque on the particle.
${\textit{Sg}}$ limits. We also modify the rotation-induced lift model of linear shear flow to account for the parabolic shear effect, which causes lift enhancement for the leading particle and lift attenuation for the lagging particle in the Poiseuille flow, compared to the linear shear case. This lift attenuation may give rise to an inverse Magnus force at low slip Reynolds numbers. In addition, the model for the particle free rotation rate for the Poiseuille flow is established by correcting the one for the linear shear flow, providing a more accurate prediction of the torque on the particle.





Pedestal origin and extrapolation of high-density small edge-localised-modes peak parallel energy fluence in ITER and SPARC
-
- Journal:
- Journal of Plasma Physics / Volume 92 / Issue 2 / April 2026
- Published online by Cambridge University Press:
- 31 March 2026, E35
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Aerodynamic unsteadiness from a forward-facing door over a cavity at Mach 6
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 31 March 2026, A6
-
- Article
- Export citation
-
A joint experimental–computational investigation was conducted to examine the aerodynamic behaviour of a partially closed cavity model in Mach-6 flow. The model, consisting of a flat plate with a rectangular cavity and a forward-facing hinged door, resulted in a strong 500 Hz fluctuation with a 7.5
 $^\circ$ door and 25 mm cavity depth. The experiments revealed a recirculation bubble present upstream of the cavity region. The fluctuations, detected by surface pressure sensors on the upper surface, upstream cavity wall and cavity floor, were caused by oscillations of the separation bubble along the streamwise axis. Notably, this phenomenon is not explained by established empirical models for cavity flows, such as the Rossiter mechanism or closed-box acoustic resonance. To further elucidate the flow physics, detached eddy simulations (DESs) of the flow were conducted, providing a detailed understanding of the complex flow phenomena. The DES results complemented the experimental data, offering insights into the unsteady flow behaviour and the mechanisms driving the pressure fluctuations. Additional experiments and simulations were conducted for other door angles to simulate different stages of opening. The strong pressure fluctuations at approximately 500 Hz were only experimentally observed for door angles between 5.0
$^\circ$ door and 25 mm cavity depth. The experiments revealed a recirculation bubble present upstream of the cavity region. The fluctuations, detected by surface pressure sensors on the upper surface, upstream cavity wall and cavity floor, were caused by oscillations of the separation bubble along the streamwise axis. Notably, this phenomenon is not explained by established empirical models for cavity flows, such as the Rossiter mechanism or closed-box acoustic resonance. To further elucidate the flow physics, detached eddy simulations (DESs) of the flow were conducted, providing a detailed understanding of the complex flow phenomena. The DES results complemented the experimental data, offering insights into the unsteady flow behaviour and the mechanisms driving the pressure fluctuations. Additional experiments and simulations were conducted for other door angles to simulate different stages of opening. The strong pressure fluctuations at approximately 500 Hz were only experimentally observed for door angles between 5.0 $^\circ$ and 7.5
$^\circ$ and 7.5 $^\circ$ but were absent at much smaller and larger angles. Additionally, several cavity depths were tested, which demonstrated that a shallower cavity delayed the onset of fluctuations until a higher free-stream Reynolds number was reached. The combination of experimental and numerical results provides valuable initial data on the aerodynamic performance of a hypersonic forward-facing door over a cavity.
$^\circ$ but were absent at much smaller and larger angles. Additionally, several cavity depths were tested, which demonstrated that a shallower cavity delayed the onset of fluctuations until a higher free-stream Reynolds number was reached. The combination of experimental and numerical results provides valuable initial data on the aerodynamic performance of a hypersonic forward-facing door over a cavity.




Staggered versus aligned: how wavy wall topology governs hypersonic boundary layer transition
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 30 March 2026, A1
-
- Article
- Export citation

Nonlinear subcritical free-surface flow past topography: exponential-asymptotics solution
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 30 March 2026, A5
-
- Article
- Export citation
-
A theoretical study is made of steady, subcritical (Froude number
 $F \lt 1$) two-dimensional free-surface flow due to a uniform stream flowing over smooth, locally confined bottom topography of large horizontal extent (
$F \lt 1$) two-dimensional free-surface flow due to a uniform stream flowing over smooth, locally confined bottom topography of large horizontal extent (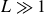 $L \gg 1$) and finite peak height (
$L \gg 1$) and finite peak height ( $\varepsilon = O(1)$). In earlier work, this flow was analysed based on the nonlinear shallow-water equations which neglect the effects of dispersion altogether. This so-called hydraulic theory predicts a steady disturbance confined in the vicinity of the topography if
$\varepsilon = O(1)$). In earlier work, this flow was analysed based on the nonlinear shallow-water equations which neglect the effects of dispersion altogether. This so-called hydraulic theory predicts a steady disturbance confined in the vicinity of the topography if  $\varepsilon$ is below a critical value
$\varepsilon$ is below a critical value  $\varepsilon _{\textit{crit}}(F)$. The present asymptotic analysis of the full potential flow equations focuses on how dispersive effects (controlled by
$\varepsilon _{\textit{crit}}(F)$. The present asymptotic analysis of the full potential flow equations focuses on how dispersive effects (controlled by 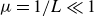 $\mu = 1/L \ll 1$) influence this steady state, particularly in regard to a steady short-scale radiating wave downstream that is ignored by hydraulic theory. Utilizing exponential asymptotics, it is shown that as
$\mu = 1/L \ll 1$) influence this steady state, particularly in regard to a steady short-scale radiating wave downstream that is ignored by hydraulic theory. Utilizing exponential asymptotics, it is shown that as  $\varepsilon$ is increased this dispersive wave, whose amplitude is formally exponentially small with respect to
$\varepsilon$ is increased this dispersive wave, whose amplitude is formally exponentially small with respect to  $\mu$, grows sharply and ultimately it becomes comparable with the hydraulic wave disturbance when
$\mu$, grows sharply and ultimately it becomes comparable with the hydraulic wave disturbance when  $\varepsilon$ approaches
$\varepsilon$ approaches  $\varepsilon _{\textit{crit}}$. Thus, the nonlinear shallow-water equations break down in the vicinity of
$\varepsilon _{\textit{crit}}$. Thus, the nonlinear shallow-water equations break down in the vicinity of  $\varepsilon _{\textit{crit}}$ regardless of
$\varepsilon _{\textit{crit}}$ regardless of 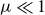 $\mu \ll 1$. The asymptotic results are supported by numerical solutions of the full potential flow theory, which also reveal a limiting
$\mu \ll 1$. The asymptotic results are supported by numerical solutions of the full potential flow theory, which also reveal a limiting  $\varepsilon$,
$\varepsilon$, 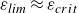 $\varepsilon _{\textit{lim}} \approx \varepsilon _{{ crit}}$, above which steady wave responses cannot be computed. For
$\varepsilon _{\textit{lim}} \approx \varepsilon _{{ crit}}$, above which steady wave responses cannot be computed. For  $\varepsilon$ just below
$\varepsilon$ just below  $\varepsilon _{\textit{lim}}$, the downstream wave resembles a steep steady Stokes periodic wave, while for
$\varepsilon _{\textit{lim}}$, the downstream wave resembles a steep steady Stokes periodic wave, while for  $\varepsilon$ slightly above
$\varepsilon$ slightly above 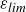 $\varepsilon _{\textit{lim}}$ unsteady computations suggest that the downstream disturbance steepens and breaks.
$\varepsilon _{\textit{lim}}$ unsteady computations suggest that the downstream disturbance steepens and breaks.

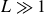



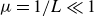





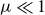

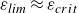



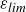

Scale-by-scale energy transfers in bubbly flows
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 30 March 2026, A12
-
- Article
- Export citation

Maxwell–Cattaneo double-diffusive convection: the general case
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 30 March 2026, A34
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Modulation induced by very-large-scale motions on the inclination angle of wall-attached eddies: an atmospheric surface layer experiment
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 30 March 2026, A24
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The forward leaning inclination angle,
 $\gamma$, of coherent turbulent structures is a well-known feature of wall-bounded turbulent flows. Although invariant across friction Reynolds numbers within the range
$\gamma$, of coherent turbulent structures is a well-known feature of wall-bounded turbulent flows. Although invariant across friction Reynolds numbers within the range  $\textit{Re}_\tau =10^3{-}10^6$,
$\textit{Re}_\tau =10^3{-}10^6$, 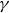 $\gamma$ can vary significantly across turbulent scales within a high-Reynolds-number flow. Very-large-scale motions (VLSMs) are known to induce significant changes in the instantaneous shear profile, which is a conditioning event that could trigger variability in the inclination angle of smaller coherent turbulent structures. Although this aspect has been extensively studied via numerical and laboratory experiments, few studies have explored this feature for a very-high-Reynolds-number atmospheric flow. In this work, the inclination angle of turbulent structures within the atmospheric surface layer at a very high Reynolds number (
$\gamma$ can vary significantly across turbulent scales within a high-Reynolds-number flow. Very-large-scale motions (VLSMs) are known to induce significant changes in the instantaneous shear profile, which is a conditioning event that could trigger variability in the inclination angle of smaller coherent turbulent structures. Although this aspect has been extensively studied via numerical and laboratory experiments, few studies have explored this feature for a very-high-Reynolds-number atmospheric flow. In this work, the inclination angle of turbulent structures within the atmospheric surface layer at a very high Reynolds number (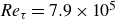 $\textit{Re}_\tau =7.9\times 10^5$) is investigated by deploying a scanning Doppler light detection and ranging and a super large particle image velocimetry (SLPIV) apparatus. The inclination angle of wall-attached eddies is inferred either from the two-point correlation of streamwise velocity (
$\textit{Re}_\tau =7.9\times 10^5$) is investigated by deploying a scanning Doppler light detection and ranging and a super large particle image velocimetry (SLPIV) apparatus. The inclination angle of wall-attached eddies is inferred either from the two-point correlation of streamwise velocity (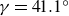 $\gamma =41.1^\circ$) or with a scale-dependent approach through the spectral linear stochastic estimator (SLSE). The SLSE (and, thus, the scale-dependent inclination angle) is conditionally evaluated based on the high- and low-momentum events induced by VLSMs, both in the streamwise (
$\gamma =41.1^\circ$) or with a scale-dependent approach through the spectral linear stochastic estimator (SLSE). The SLSE (and, thus, the scale-dependent inclination angle) is conditionally evaluated based on the high- and low-momentum events induced by VLSMs, both in the streamwise ( $u'_{\textit{VLSM}}$) and in the vertical (
$u'_{\textit{VLSM}}$) and in the vertical ( $w'_{\textit{VLSM}}$) velocity components. As a result, lower inclination angles (
$w'_{\textit{VLSM}}$) velocity components. As a result, lower inclination angles (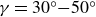 $\gamma =30^\circ {-}50^\circ$) are found for
$\gamma =30^\circ {-}50^\circ$) are found for 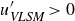 $u'_{\textit{VLSM}}\gt 0$ (
$u'_{\textit{VLSM}}\gt 0$ ( $w'_{\textit{VLSM}}\lt 0$), while higher values (
$w'_{\textit{VLSM}}\lt 0$), while higher values ( $50^\circ {-}85^\circ$) are ascribed to
$50^\circ {-}85^\circ$) are ascribed to 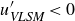 $u'_{\textit{VLSM}}\lt 0$ (
$u'_{\textit{VLSM}}\lt 0$ (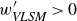 $w'_{\textit{VLSM}}\gt 0$). This result emphasises the primary role that VLSMs play in shaping the wall-attached eddy geometry, which, in turn, is crucial to determine the Reynolds stress balance within the wall-attached eddy range.
$w'_{\textit{VLSM}}\gt 0$). This result emphasises the primary role that VLSMs play in shaping the wall-attached eddy geometry, which, in turn, is crucial to determine the Reynolds stress balance within the wall-attached eddy range.


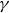
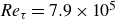
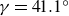


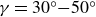
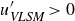


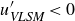
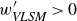

Controlled particle displacement by hydrodynamic obstacle interaction in non-inertial flows
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 30 March 2026, A16
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Transport structure of two-dimensional homogeneous Rayleigh–Bénard convection
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 30 March 2026, A11
-
- Article
- Export citation

The role of Lagrangian drift in the generation of surface waves by wind
-
- Journal:
- Journal of Fluid Mechanics / Volume 1032 / 10 April 2026
- Published online by Cambridge University Press:
- 30 March 2026, A9
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
