Refine listing
Actions for selected content:
593 results in 35-XX
A SIMPLE AND NEARLY OPTIMAL INVESTMENT STRATEGY TO MINIMIZE THE PROBABILITY OF LIFETIME RUIN
- Part of
-
- Journal:
- ASTIN Bulletin: The Journal of the IAA / Volume 52 / Issue 2 / May 2022
- Published online by Cambridge University Press:
- 16 February 2022, pp. 619-643
- Print publication:
- May 2022
-
- Article
- Export citation
Bifurcation of elastic curves with modulated stiffness
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 34 / Issue 1 / February 2023
- Published online by Cambridge University Press:
- 28 January 2022, pp. 28-54
-
- Article
-
- You have access
- HTML
- Export citation
Necessary and sufficient conditions on global solvability for the p-k-Hessian inequalities
- Part of
-
- Journal:
- Canadian Mathematical Bulletin / Volume 65 / Issue 4 / December 2022
- Published online by Cambridge University Press:
- 17 January 2022, pp. 1004-1019
- Print publication:
- December 2022
-
- Article
- Export citation
-
In this paper, we discuss the solvability of the p-k-Hessian inequality
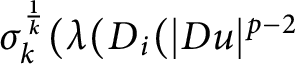 $\sigma _{k}^{\frac 1k} ( \lambda ( D_{i} (|Du|^{p-2}$
$\sigma _{k}^{\frac 1k} ( \lambda ( D_{i} (|Du|^{p-2}$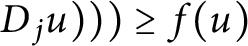 $ D_{j}u ) ) ) \geq f(u)$ on the entire space
$ D_{j}u ) ) ) \geq f(u)$ on the entire space 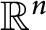 $\mathbb {R}^{n}$ and provide a necessary and sufficient condition, which can be regarded as a generalized Keller–Osserman condition. Furthermore, we obtain the optimal regularity of solution.
$\mathbb {R}^{n}$ and provide a necessary and sufficient condition, which can be regarded as a generalized Keller–Osserman condition. Furthermore, we obtain the optimal regularity of solution.
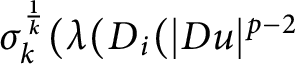
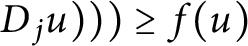
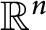
Unconditional uniqueness for the energy-critical nonlinear Schrödinger equation on
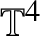 $\mathbb {T}^{4}$
$\mathbb {T}^{4}$
- Part of
-
- Journal:
- Forum of Mathematics, Pi / Volume 10 / 2022
- Published online by Cambridge University Press:
- 13 January 2022, e3
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider the
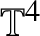 $\mathbb {T}^{4}$ cubic nonlinear Schrödinger equation (NLS), which is energy-critical. We study the unconditional uniqueness of solutions to the NLS via the cubic Gross–Pitaevskii hierarchy, an uncommon method for NLS analysis which is being explored [24, 35] and does not require the existence of a solution in Strichartz-type spaces. We prove U-V multilinear estimates to replace the previously used Sobolev multilinear estimates. To incorporate the weaker estimates, we work out new combinatorics from scratch and compute, for the first time, the time integration limits, in the recombined Duhamel–Born expansion. The new combinatorics and the U-V estimates then seamlessly conclude the
$\mathbb {T}^{4}$ cubic nonlinear Schrödinger equation (NLS), which is energy-critical. We study the unconditional uniqueness of solutions to the NLS via the cubic Gross–Pitaevskii hierarchy, an uncommon method for NLS analysis which is being explored [24, 35] and does not require the existence of a solution in Strichartz-type spaces. We prove U-V multilinear estimates to replace the previously used Sobolev multilinear estimates. To incorporate the weaker estimates, we work out new combinatorics from scratch and compute, for the first time, the time integration limits, in the recombined Duhamel–Born expansion. The new combinatorics and the U-V estimates then seamlessly conclude the 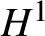 $H^{1}$ unconditional uniqueness for the NLS under the infinite-hierarchy framework. This work establishes a unified scheme to prove
$H^{1}$ unconditional uniqueness for the NLS under the infinite-hierarchy framework. This work establishes a unified scheme to prove 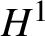 $H^{1}$ uniqueness for the
$H^{1}$ uniqueness for the 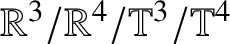 $ \mathbb {R}^{3}/\mathbb {R}^{4}/\mathbb {T}^{3}/\mathbb {T}^{4}$ energy-critical Gross–Pitaevskii hierarchies and thus the corresponding NLS.
$ \mathbb {R}^{3}/\mathbb {R}^{4}/\mathbb {T}^{3}/\mathbb {T}^{4}$ energy-critical Gross–Pitaevskii hierarchies and thus the corresponding NLS.
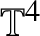
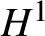
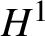
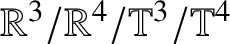
Periodic solutions for one-dimensional nonlinear nonlocal problem with drift including singular nonlinearities
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 153 / Issue 1 / February 2023
- Published online by Cambridge University Press:
- 23 December 2021, pp. 229-261
- Print publication:
- February 2023
-
- Article
- Export citation
-
In this paper, we prove existence results of a one-dimensional periodic solution to equations with the fractional Laplacian of order $s\in (1/2,1)$
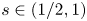 , singular nonlinearity and gradient term under various situations, including nonlocal contra-part of classical Lienard vector equations, as well other nonlocal versions of classical results know only in the context of second-order ODE. Our proofs are based on degree theory and Perron's method, so before that we need to establish a variety of priori estimates under different assumptions on the nonlinearities appearing in the equations. Besides, we obtain also multiplicity results in a regime where a priori bounds are lost and bifurcation from infinity occurs.
, singular nonlinearity and gradient term under various situations, including nonlocal contra-part of classical Lienard vector equations, as well other nonlocal versions of classical results know only in the context of second-order ODE. Our proofs are based on degree theory and Perron's method, so before that we need to establish a variety of priori estimates under different assumptions on the nonlinearities appearing in the equations. Besides, we obtain also multiplicity results in a regime where a priori bounds are lost and bifurcation from infinity occurs.
Optimal Hardy-weights for the (p, A)-Laplacian with a potential term
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 153 / Issue 1 / February 2023
- Published online by Cambridge University Press:
- 17 December 2021, pp. 289-306
- Print publication:
- February 2023
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We construct new optimal
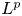 $L^{p}$ Hardy-type inequalities for elliptic Schrödinger-type operators with a potential term.
$L^{p}$ Hardy-type inequalities for elliptic Schrödinger-type operators with a potential term.
The fragmentation equation with size diffusion: Well posedness and long-term behaviour
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 33 / Issue 6 / December 2022
- Published online by Cambridge University Press:
- 16 December 2021, pp. 1083-1116
-
- Article
- Export citation
-
The dynamics of the fragmentation equation with size diffusion is investigated when the size ranges in
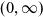 $(0,\infty)$. The associated linear operator involves three terms and can be seen as a nonlocal perturbation of a Schrödinger operator. A Miyadera perturbation argument is used to prove that it is the generator of a positive, analytic semigroup on a weighted
$(0,\infty)$. The associated linear operator involves three terms and can be seen as a nonlocal perturbation of a Schrödinger operator. A Miyadera perturbation argument is used to prove that it is the generator of a positive, analytic semigroup on a weighted 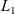 $L_1$-space. Moreover, if the overall fragmentation rate does not vanish at infinity, then there is a unique stationary solution with given mass. Assuming further that the overall fragmentation rate diverges to infinity for large sizes implies the immediate compactness of the semigroup and that it eventually stabilizes at an exponential rate to a one-dimensional projection carrying the information of the mass of the initial value.
$L_1$-space. Moreover, if the overall fragmentation rate does not vanish at infinity, then there is a unique stationary solution with given mass. Assuming further that the overall fragmentation rate diverges to infinity for large sizes implies the immediate compactness of the semigroup and that it eventually stabilizes at an exponential rate to a one-dimensional projection carrying the information of the mass of the initial value.
A sharpened form of Adams-type inequalities on higher-order Sobolev spaces
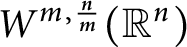 $W^{m,\frac {n}{m}}(\mathbb {R}^n)$: a simple approach
$W^{m,\frac {n}{m}}(\mathbb {R}^n)$: a simple approach
- Part of
-
- Journal:
- Canadian Mathematical Bulletin / Volume 65 / Issue 4 / December 2022
- Published online by Cambridge University Press:
- 15 December 2021, pp. 895-905
- Print publication:
- December 2022
-
- Article
- Export citation
-
In this paper, we develop an extremely simple method to establish the sharpened Adams-type inequalities on higher-order Sobolev spaces
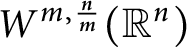 $W^{m,\frac {n}{m}}(\mathbb {R}^n)$ in the entire space
$W^{m,\frac {n}{m}}(\mathbb {R}^n)$ in the entire space 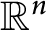 $\mathbb {R}^n$, which can be stated as follows: Given
$\mathbb {R}^n$, which can be stated as follows: Given 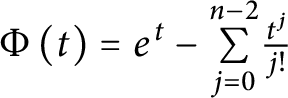 $\Phi \left ( t\right ) =e^{t}-\underset {j=0}{\overset {n-2}{\sum }} \frac {t^{j}}{j!}$ and the Adams sharp constant
$\Phi \left ( t\right ) =e^{t}-\underset {j=0}{\overset {n-2}{\sum }} \frac {t^{j}}{j!}$ and the Adams sharp constant 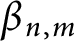 $\beta _{n,m}$. Then, for any
$\beta _{n,m}$. Then, for any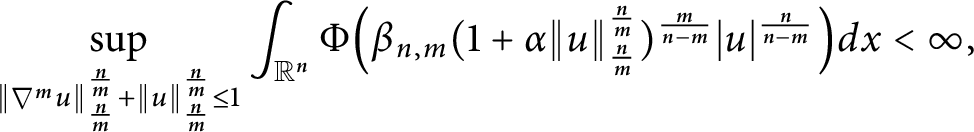 $$ \begin{align*}\sup_{\|\nabla^mu\|_{\frac{n}{m}}^{\frac{n}{m}}+\|u\|_{\frac{n}{m}}^{\frac{n}{m}}\leq1}\int_{\mathbb{R}^n}\Phi\Big(\beta_{n,m} (1+\alpha \|u\|_{\frac{n}{m}}^{\frac{n}{m}} )^{\frac{m}{n-m}}|u|^{\frac{n}{n-m}}\Big)dx<\infty, \end{align*} $$
$$ \begin{align*}\sup_{\|\nabla^mu\|_{\frac{n}{m}}^{\frac{n}{m}}+\|u\|_{\frac{n}{m}}^{\frac{n}{m}}\leq1}\int_{\mathbb{R}^n}\Phi\Big(\beta_{n,m} (1+\alpha \|u\|_{\frac{n}{m}}^{\frac{n}{m}} )^{\frac{m}{n-m}}|u|^{\frac{n}{n-m}}\Big)dx<\infty, \end{align*} $$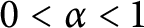 $0<\alpha <1$. Furthermore, we construct a proper test function sequence to derive the sharpness of the exponent
$0<\alpha <1$. Furthermore, we construct a proper test function sequence to derive the sharpness of the exponent 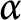 $\alpha $ of the above Adams inequalities. Namely, we will show that if
$\alpha $ of the above Adams inequalities. Namely, we will show that if 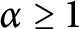 $\alpha \ge 1$, then the above supremum is infinite.
$\alpha \ge 1$, then the above supremum is infinite.Our argument avoids applying the complicated blow-up analysis often used in the literature to deal with such sharpened inequalities.
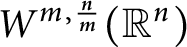
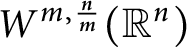
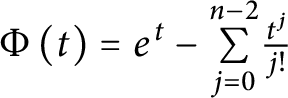
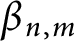
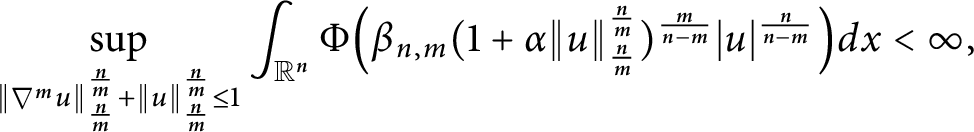
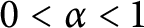
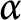
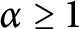
Liouville-type results for positive solutions of pseudo-relativistic Schrödinger system
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 153 / Issue 1 / February 2023
- Published online by Cambridge University Press:
- 13 December 2021, pp. 196-228
- Print publication:
- February 2023
-
- Article
- Export citation
-
In this paper, we are concerned with the physically engaging pseudo-relativistic Schrödinger system:
\[ \begin{cases} \left(-\Delta+m^{2}\right)^{s}u(x)=f(x,u,v,\nabla u) & \hbox{in } \Omega,\\ \left(-\Delta+m^{2}\right)^{t}v(x)=g(x,u,v,\nabla v) & \hbox{in } \Omega,\\ u>0,v>0 & \hbox{in } \Omega, \\ u=v\equiv 0 & \hbox{in } \mathbb{R}^{N}\setminus\Omega, \end{cases} \]where $s,t\in (0,1)$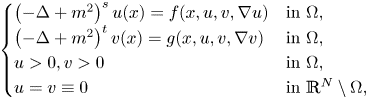
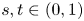 and the mass $m>0.$
and the mass $m>0.$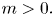 By using the direct method of moving plane, we prove the strict monotonicity, symmetry and uniqueness for positive solutions to the above system in a bounded domain, unbounded domain, $\mathbb {R}^{N}$
By using the direct method of moving plane, we prove the strict monotonicity, symmetry and uniqueness for positive solutions to the above system in a bounded domain, unbounded domain, $\mathbb {R}^{N}$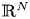 , $\mathbb {R}^{N}_{+}$
, $\mathbb {R}^{N}_{+}$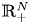 and a coercive epigraph domain $\Omega$
and a coercive epigraph domain $\Omega$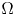 in $\mathbb {R}^{N}$
in $\mathbb {R}^{N}$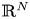 , respectively.
, respectively.
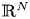
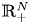
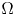
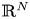
Decay of weak solutions to Vlasov equation coupled with a shear thickening fluid
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 153 / Issue 1 / February 2023
- Published online by Cambridge University Press:
- 02 December 2021, pp. 167-176
- Print publication:
- February 2023
-
- Article
- Export citation
-
We show that the energy norm of weak solutions to Vlasov equation coupled with a shear thickening fluid on the whole space has a decay rate the energy norm $E(t) \leq {C}/{(1+t)^{\alpha }}, \forall t \geq 0$
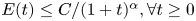 for $\alpha \in (0,3/2)$
for $\alpha \in (0,3/2)$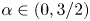 .
.
Partial regularity for minimizers of discontinuous quasiconvex integrals with general growth
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 152 / Issue 5 / October 2022
- Published online by Cambridge University Press:
- 01 December 2021, pp. 1191-1232
- Print publication:
- October 2022
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We prove the partial Hölder continuity for minimizers of quasiconvex functionals
where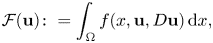 \begin{equation*} \mathcal{F}({\bf u}) \colon =\int_{\Omega} f(x,{\bf u},D{\bf u})\,\textrm{d} x, \end{equation*}
\begin{equation*} \mathcal{F}({\bf u}) \colon =\int_{\Omega} f(x,{\bf u},D{\bf u})\,\textrm{d} x, \end{equation*}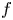 $f$ satisfies a uniform VMO condition with respect to the
$f$ satisfies a uniform VMO condition with respect to the 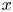 $x$-variable and is continuous with respect to
$x$-variable and is continuous with respect to 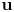 ${\bf u}$. The growth condition with respect to the gradient variable is assumed a general one.
${\bf u}$. The growth condition with respect to the gradient variable is assumed a general one.
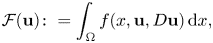
A weight-homogenous condition to the real Jacobian conjecture in $ {\mathbb {R}}^{2}$
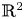
- Part of
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 64 / Issue 4 / November 2021
- Published online by Cambridge University Press:
- 19 November 2021, pp. 1028-1036
-
- Article
- Export citation
-
It is known that a polynomial local diffeomorphism $(f,\, g): {\mathbb {R}}^{2} \to {\mathbb {R}}^{2}$
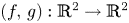 is a global diffeomorphism provided the higher homogeneous terms of $f f_x+g g_x$
is a global diffeomorphism provided the higher homogeneous terms of $f f_x+g g_x$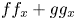 and $f f_y+g g_y$
and $f f_y+g g_y$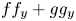 do not have real linear factors in common. Here, we give a weight-homogeneous framework of this result. Our approach uses qualitative theory of differential equations. In our reasoning, we obtain a result on polynomial Hamiltonian vector fields in the plane, generalization of a known fact.
do not have real linear factors in common. Here, we give a weight-homogeneous framework of this result. Our approach uses qualitative theory of differential equations. In our reasoning, we obtain a result on polynomial Hamiltonian vector fields in the plane, generalization of a known fact.
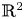
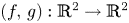
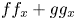
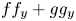
Existence and regularity of time-dependent pullback attractors for the non-autonomous nonclassical diffusion equations
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 152 / Issue 6 / December 2022
- Published online by Cambridge University Press:
- 18 November 2021, pp. 1533-1550
- Print publication:
- December 2022
-
- Article
- Export citation
-
In this paper, we prove the existence and regularity of pullback attractors for non-autonomous nonclassical diffusion equations with nonlocal diffusion when the nonlinear term satisfies critical exponential growth and the external force term
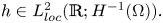 $h \in L_{l o c}^{2}(\mathbb {R} ; H^{-1}(\Omega )).$ Under some appropriate assumptions, we establish the existence and uniqueness of the weak solution in the time-dependent space
$h \in L_{l o c}^{2}(\mathbb {R} ; H^{-1}(\Omega )).$ Under some appropriate assumptions, we establish the existence and uniqueness of the weak solution in the time-dependent space 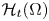 $\mathcal {H}_{t}(\Omega )$ and the existence and regularity of the pullback attractors.
$\mathcal {H}_{t}(\Omega )$ and the existence and regularity of the pullback attractors.
On the global existence and qualitative behaviour of one-dimensional solutions to a model for urban crime
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 33 / Issue 5 / October 2022
- Published online by Cambridge University Press:
- 02 November 2021, pp. 919-959
-
- Article
- Export citation
-
We consider the no-flux initial-boundary value problem for the cross-diffusive evolution system:
which was introduced by Short et al. in [40] with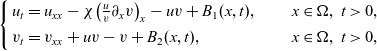 \begin{eqnarray*}\left\{ \begin{array}{ll}u_t = u_{xx} - \chi \big(\frac{u}{v} \partial_x v \big)_x - uv +B_1(x,t),\qquad & x\in \Omega, \ t>0, \\[1mm]v_t = v_{xx} +uv - v + B_2(x,t),\qquad & x\in \Omega, \ t>0,\end{array} \right.\end{eqnarray*}
\begin{eqnarray*}\left\{ \begin{array}{ll}u_t = u_{xx} - \chi \big(\frac{u}{v} \partial_x v \big)_x - uv +B_1(x,t),\qquad & x\in \Omega, \ t>0, \\[1mm]v_t = v_{xx} +uv - v + B_2(x,t),\qquad & x\in \Omega, \ t>0,\end{array} \right.\end{eqnarray*}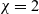 $\chi=2$ to describe the dynamics of urban crime.
$\chi=2$ to describe the dynamics of urban crime.In bounded intervals
 $\Omega\subset\mathbb{R}$ and with prescribed suitably regular non-negative functions
$\Omega\subset\mathbb{R}$ and with prescribed suitably regular non-negative functions  $B_1$ and
$B_1$ and 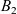 $B_2$, we first prove the existence of global classical solutions for any choice of
$B_2$, we first prove the existence of global classical solutions for any choice of 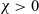 $\chi>0$ and all reasonably regular non-negative initial data.
$\chi>0$ and all reasonably regular non-negative initial data.We next address the issue of determining the qualitative behaviour of solutions under appropriate assumptions on the asymptotic properties of
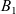 $B_1$ and
$B_1$ and 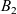 $B_2$. Indeed, for arbitrary
$B_2$. Indeed, for arbitrary 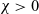 $\chi>0$, we obtain boundedness of the solutions given strict positivity of the average of
$\chi>0$, we obtain boundedness of the solutions given strict positivity of the average of 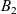 $B_2$ over the domain; moreover, it is seen that imposing a mild decay assumption on
$B_2$ over the domain; moreover, it is seen that imposing a mild decay assumption on 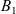 $B_1$ implies that u must decay to zero in the long-term limit. Our final result, valid for all
$B_1$ implies that u must decay to zero in the long-term limit. Our final result, valid for all 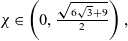 $\chi\in\left(0,\frac{\sqrt{6\sqrt{3}+9}}{2}\right),$ which contains the relevant value
$\chi\in\left(0,\frac{\sqrt{6\sqrt{3}+9}}{2}\right),$ which contains the relevant value 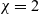 $\chi=2$, states that under the above decay assumption on
$\chi=2$, states that under the above decay assumption on 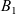 $B_1$, if furthermore
$B_1$, if furthermore 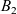 $B_2$ appropriately stabilises to a non-trivial function
$B_2$ appropriately stabilises to a non-trivial function 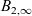 $B_{2,\infty}$, then (u,v) approaches the limit
$B_{2,\infty}$, then (u,v) approaches the limit 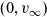 $(0,v_\infty)$, where
$(0,v_\infty)$, where 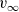 $v_\infty$ denotes the solution of We conclude with some numerical simulations exploring possible effects that may arise when considering large values of
$v_\infty$ denotes the solution of We conclude with some numerical simulations exploring possible effects that may arise when considering large values of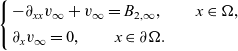 \begin{eqnarray*}\left\{ \begin{array}{l}-\partial_{xx}v_\infty + v_\infty = B_{2,\infty},\qquad x\in \Omega, \\[1mm]\partial_x v_{\infty}=0,\qquad x\in\partial\Omega.\end{array} \right.\end{eqnarray*}
\begin{eqnarray*}\left\{ \begin{array}{l}-\partial_{xx}v_\infty + v_\infty = B_{2,\infty},\qquad x\in \Omega, \\[1mm]\partial_x v_{\infty}=0,\qquad x\in\partial\Omega.\end{array} \right.\end{eqnarray*}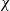 $\chi$ not covered by our qualitative analysis. We observe that when
$\chi$ not covered by our qualitative analysis. We observe that when 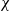 $\chi$ increases, solutions may grow substantially on short time intervals, whereas only on large timescales diffusion will dominate and enforce equilibration.
$\chi$ increases, solutions may grow substantially on short time intervals, whereas only on large timescales diffusion will dominate and enforce equilibration.
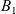
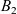
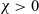
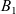
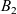
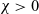
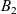
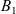
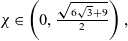
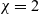
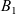
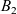
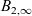
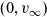
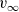
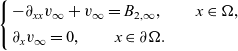
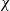
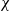
BOUNDS ON THE CRITICAL TIMES FOR THE GENERAL FISHER–KPP EQUATION
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 63 / Issue 4 / October 2021
- Published online by Cambridge University Press:
- 02 November 2021, pp. 448-468
-
- Article
- Export citation
Charge transport modelling of Lithium-ion batteries
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 33 / Issue 6 / December 2022
- Published online by Cambridge University Press:
- 21 October 2021, pp. 983-1031
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Existence results for a nonlinear nonautonomous transmission problem via domain perturbation
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 152 / Issue 6 / December 2022
- Published online by Cambridge University Press:
- 11 October 2021, pp. 1451-1476
- Print publication:
- December 2022
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In this paper we study the existence and the analytic dependence upon domain perturbation of the solutions of a nonlinear nonautonomous transmission problem for the Laplace equation. The problem is defined in a pair of sets consisting of a perforated domain and an inclusion whose shape is determined by a suitable diffeomorphism
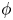 $\phi$. First we analyse the case in which the inclusion is a fixed domain. Then we will perturb the inclusion and study the arising boundary value problem and the dependence of a specific family of solutions upon the perturbation parameter
$\phi$. First we analyse the case in which the inclusion is a fixed domain. Then we will perturb the inclusion and study the arising boundary value problem and the dependence of a specific family of solutions upon the perturbation parameter 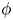 $\phi$.
$\phi$.
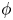
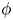
Lp-trace-free generalized Korn inequalities for incompatible tensor fields in three space dimensions
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 152 / Issue 6 / December 2022
- Published online by Cambridge University Press:
- 11 October 2021, pp. 1477-1508
- Print publication:
- December 2022
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
For
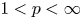 $1< p<\infty$ we prove an
$1< p<\infty$ we prove an 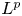 $L^{p}$-version of the generalized trace-free Korn inequality for incompatible tensor fields
$L^{p}$-version of the generalized trace-free Korn inequality for incompatible tensor fields 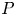 $P$ in
$P$ in 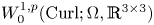 $W^{1,p}_0(\operatorname {Curl}; \Omega ,\mathbb {R}^{3\times 3})$. More precisely, let
$W^{1,p}_0(\operatorname {Curl}; \Omega ,\mathbb {R}^{3\times 3})$. More precisely, let 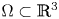 $\Omega \subset \mathbb {R}^{3}$ be a bounded Lipschitz domain. Then there exists a constant
$\Omega \subset \mathbb {R}^{3}$ be a bounded Lipschitz domain. Then there exists a constant 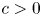 $c>0$ such thatholds for all tensor fields
$c>0$ such thatholds for all tensor fields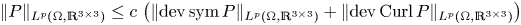 \[ \lVert{ P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}\leq c\,\left(\lVert{\operatorname{dev} \operatorname{sym} P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})} + \lVert{ \operatorname{dev} \operatorname{Curl} P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}\right) \]
\[ \lVert{ P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}\leq c\,\left(\lVert{\operatorname{dev} \operatorname{sym} P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})} + \lVert{ \operatorname{dev} \operatorname{Curl} P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}\right) \]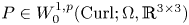 $P\in W^{1,p}_0(\operatorname {Curl}; \Omega ,\mathbb {R}^{3\times 3})$, i.e., for all
$P\in W^{1,p}_0(\operatorname {Curl}; \Omega ,\mathbb {R}^{3\times 3})$, i.e., for all 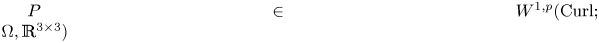 $P\in W^{1,p} (\operatorname {Curl}; \Omega ,\mathbb {R}^{3\times 3})$ with vanishing tangential trace
$P\in W^{1,p} (\operatorname {Curl}; \Omega ,\mathbb {R}^{3\times 3})$ with vanishing tangential trace 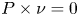 $P\times \nu =0$ on
$P\times \nu =0$ on 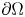 $\partial \Omega$ where
$\partial \Omega$ where 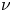 $\nu$ denotes the outward unit normal vector field to
$\nu$ denotes the outward unit normal vector field to 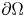 $\partial \Omega$ and
$\partial \Omega$ and 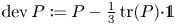 $\operatorname {dev} P : = P -\frac 13 \operatorname {tr}(P) {\cdot } {\mathbb {1}}$ denotes the deviatoric (trace-free) part of
$\operatorname {dev} P : = P -\frac 13 \operatorname {tr}(P) {\cdot } {\mathbb {1}}$ denotes the deviatoric (trace-free) part of 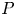 $P$. We also show the norm equivalencefor tensor fields
$P$. We also show the norm equivalencefor tensor fields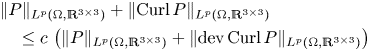 \begin{align*} &\lVert{ P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}+\lVert{ \operatorname{Curl} P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}\\ &\quad\leq c\,\left(\lVert{P}\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})} + \lVert{ \operatorname{dev} \operatorname{Curl} P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}\right) \end{align*}
\begin{align*} &\lVert{ P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}+\lVert{ \operatorname{Curl} P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}\\ &\quad\leq c\,\left(\lVert{P}\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})} + \lVert{ \operatorname{dev} \operatorname{Curl} P }\rVert_{L^{p}(\Omega,\mathbb{R}^{3\times 3})}\right) \end{align*}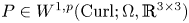 $P\in W^{1,p}(\operatorname {Curl}; \Omega ,\mathbb {R}^{3\times 3})$. These estimates also hold true for tensor fields with vanishing tangential trace only on a relatively open (non-empty) subset
$P\in W^{1,p}(\operatorname {Curl}; \Omega ,\mathbb {R}^{3\times 3})$. These estimates also hold true for tensor fields with vanishing tangential trace only on a relatively open (non-empty) subset 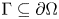 $\Gamma \subseteq \partial \Omega$ of the boundary.
$\Gamma \subseteq \partial \Omega$ of the boundary.
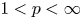
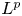
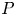
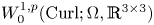
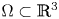
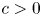
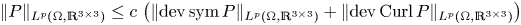
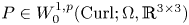
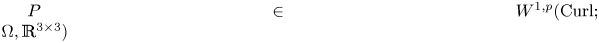
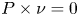
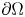
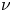
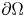
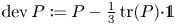
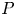
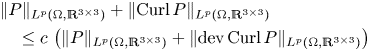
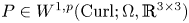
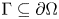
Decay properties for the incompressible Navier-Stokes flows in a half space
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics / Volume 152 / Issue 6 / December 2022
- Published online by Cambridge University Press:
- 11 October 2021, pp. 1509-1532
- Print publication:
- December 2022
-
- Article
- Export citation
-
In this article, we give a comprehensive characterization of
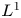 $L^1$-summability for the Navier-Stokes flows in the half space, which is a long-standing problem. The main difficulties are that
$L^1$-summability for the Navier-Stokes flows in the half space, which is a long-standing problem. The main difficulties are that  $L^q-L^r$ estimates for the Stokes flow don't work in this end-point case:
$L^q-L^r$ estimates for the Stokes flow don't work in this end-point case: 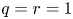 $q=r=1$; the projection operator
$q=r=1$; the projection operator 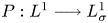 $P: L^1\longrightarrow L^1_\sigma$ is not bounded any more; useful information on the pressure function is missing, which arises in the net force exerted by the fluid on the noncompact boundary. In order to achieve our aims, by making full use of the special structure of the half space, we decompose the pressure function into two parts. Then the knotty problem of handling the pressure term can be transformed into establishing a crucial and new weighted
$P: L^1\longrightarrow L^1_\sigma$ is not bounded any more; useful information on the pressure function is missing, which arises in the net force exerted by the fluid on the noncompact boundary. In order to achieve our aims, by making full use of the special structure of the half space, we decompose the pressure function into two parts. Then the knotty problem of handling the pressure term can be transformed into establishing a crucial and new weighted 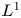 $L^1$-estimate, which plays a fundamental role. In addition, we overcome the unboundedness of the projection
$L^1$-estimate, which plays a fundamental role. In addition, we overcome the unboundedness of the projection 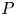 $P$ by solving an elliptic problem with homogeneous Neumann boundary condition.
$P$ by solving an elliptic problem with homogeneous Neumann boundary condition.

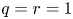
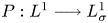
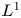
PLANAR IMMERSIONS WITH PRESCRIBED CURL AND JACOBIAN DETERMINANT ARE UNIQUE
- Part of
-
- Journal:
- Bulletin of the Australian Mathematical Society / Volume 106 / Issue 1 / August 2022
- Published online by Cambridge University Press:
- 08 October 2021, pp. 126-131
- Print publication:
- August 2022
-
- Article
- Export citation
