Refine listing
Actions for selected content:
1705 results in 60Jxx
Computable Bounds on the Spectral Gap for Unreliable Jackson Networks
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 2 / June 2015
- Published online by Cambridge University Press:
- 22 February 2016, pp. 402-424
- Print publication:
- June 2015
-
- Article
-
- You have access
- Export citation
A Recursion Formula for the Moments of the First Passage Time of the Ornstein-Uhlenbeck Process
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 2 / June 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 595-601
- Print publication:
- June 2015
-
- Article
-
- You have access
- Export citation
On Binomial Observations of Continuous-Time Markovian Population Models
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 2 / June 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 457-472
- Print publication:
- June 2015
-
- Article
-
- You have access
- Export citation
Exponential Growth of Bifurcating Processes with Ancestral Dependence
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 2 / June 2015
- Published online by Cambridge University Press:
- 22 February 2016, pp. 545-564
- Print publication:
- June 2015
-
- Article
-
- You have access
- Export citation
New Results on a Generalized Coupon Collector Problem Using Markov Chains
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 2 / June 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 405-418
- Print publication:
- June 2015
-
- Article
-
- You have access
- Export citation
The Coalescent in Peripatric Metapopulations
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 2 / June 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 538-557
- Print publication:
- June 2015
-
- Article
-
- You have access
- Export citation
Skeletons of Near-Critical Bienaymé-Galton-Watson Branching Processes
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 2 / June 2015
- Published online by Cambridge University Press:
- 22 February 2016, pp. 530-544
- Print publication:
- June 2015
-
- Article
-
- You have access
- Export citation
Sample-Path Optimal Stationary Policies in Stable Markov Decision Chains with the Average Reward Criterion
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 2 / June 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 419-440
- Print publication:
- June 2015
-
- Article
-
- You have access
- Export citation
SOME FELLER SEMIGROUPS ON
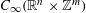 $C_{\infty }(\mathbb{R}^{n}\times \mathbb{Z}^{m})$ GENERATED BY PSEUDO-DIFFERENTIAL OPERATORS
$C_{\infty }(\mathbb{R}^{n}\times \mathbb{Z}^{m})$ GENERATED BY PSEUDO-DIFFERENTIAL OPERATORS
- Part of
-
- Journal:
- Mathematika / Volume 61 / Issue 2 / May 2015
- Published online by Cambridge University Press:
- 15 April 2015, pp. 402-413
- Print publication:
- May 2015
-
- Article
- Export citation
-
In this paper we construct some Feller semigroups, hence Feller processes, with state space
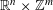 $\mathbb{R}^{n}\times \mathbb{Z}^{m}$ starting with pseudo-differential operators having symbols defined on
$\mathbb{R}^{n}\times \mathbb{Z}^{m}$ starting with pseudo-differential operators having symbols defined on 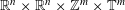 $\mathbb{R}^{n}\times \mathbb{R}^{n}\times \mathbb{Z}^{m}\times \mathbb{T}^{m}$.
$\mathbb{R}^{n}\times \mathbb{R}^{n}\times \mathbb{Z}^{m}\times \mathbb{T}^{m}$.
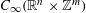
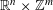
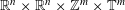
A Combinatorial Approach to a Model of Constrained Random Walkers
- Part of
-
- Journal:
- Combinatorics, Probability and Computing / Volume 25 / Issue 2 / March 2016
- Published online by Cambridge University Press:
- 16 March 2015, pp. 222-235
-
- Article
- Export citation
-
In [1], the authors consider a random walk (Z n,1, . . ., Z n,K+1) ∈
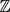 ${\mathbb{Z}}$ K+1 with the constraint that each coordinate of the walk is at distance one from the following coordinate. A functional central limit theorem for the first coordinate is proved and the limit variance is explicited. In this paper, we study an extended version of this model by conditioning the extremal coordinates to be at some fixed distance at every time. We prove a functional central limit theorem for this random walk. Using combinatorial tools, we give a precise formula of the variance and compare it with that obtained in [1].
${\mathbb{Z}}$ K+1 with the constraint that each coordinate of the walk is at distance one from the following coordinate. A functional central limit theorem for the first coordinate is proved and the limit variance is explicited. In this paper, we study an extended version of this model by conditioning the extremal coordinates to be at some fixed distance at every time. We prove a functional central limit theorem for this random walk. Using combinatorial tools, we give a precise formula of the variance and compare it with that obtained in [1].
Occupation Times, Drawdowns, and Drawups for One-Dimensional Regular Diffusions
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 04 January 2016, pp. 210-230
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
Ergodic Inequality of a Two-Parameter Infinitely-Many-Alleles Diffusion Model
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 238-246
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
A Limit Theorem for a Weiss Epidemic Process
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 247-257
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
Error Bounds for Augmented Truncations of Discrete-Time Block-Monotone Markov Chains under Geometric Drift Conditions
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 04 January 2016, pp. 83-105
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
New Results for the Two-Stage Contact Process
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 258-268
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
Impulsive Control for Continuous-Time Markov Decision Processes
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 04 January 2016, pp. 106-127
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
The Euler Scheme for a Stochastic Differential Equation Driven by Pure Jump Semimartingales
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 52 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 30 January 2018, pp. 149-166
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
BRAVO for Many-Server QED Systems with Finite Buffers
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 04 January 2016, pp. 231-250
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
Convergence in a Multidimensional Randomized Keynesian Beauty Contest
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 04 January 2016, pp. 57-82
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
Infection Spread in Random Geometric Graphs
- Part of
-
- Journal:
- Advances in Applied Probability / Volume 47 / Issue 1 / March 2015
- Published online by Cambridge University Press:
- 04 January 2016, pp. 164-181
- Print publication:
- March 2015
-
- Article
-
- You have access
- Export citation
-
In this paper we study the speed of infection spread and the survival of the contact process in the random geometric graph G = G(n, r n , f) of n nodes independently distributed in S = [-½, ½]2 according to a certain density f(·). In the first part of the paper we assume that infection spreads from one node to another at unit rate and that infected nodes stay in the same state forever. We provide an explicit lower bound on the speed of infection spread and prove that infection spreads in G with speed at least D 1 nr n 2. In the second part of the paper we consider the contact process ξt on G where infection spreads at rate λ > 0 from one node to another and each node independently recovers at unit rate. We prove that, for every λ > 0, with high probability, the contact process on G survives for an exponentially long time; there exist positive constants c 1 and c 2 such that, with probability at least 1 - c 1 / n 4, the contact process
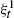 starting with all nodes infected survives up to time t n = exp(c 2 n/logn) for all n.
starting with all nodes infected survives up to time t n = exp(c 2 n/logn) for all n.
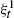 starting with all nodes infected survives up to time
starting with all nodes infected survives up to time 