Refine listing
Actions for selected content:
308 results in 35Kxx
Global boundedness and stabilization of solutions for a chemotaxis system with acceleration and logistic source
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 04 July 2025, pp. 1-30
-
- Article
- Export citation
-
This article is concerned with the following chemotaxis-growth system
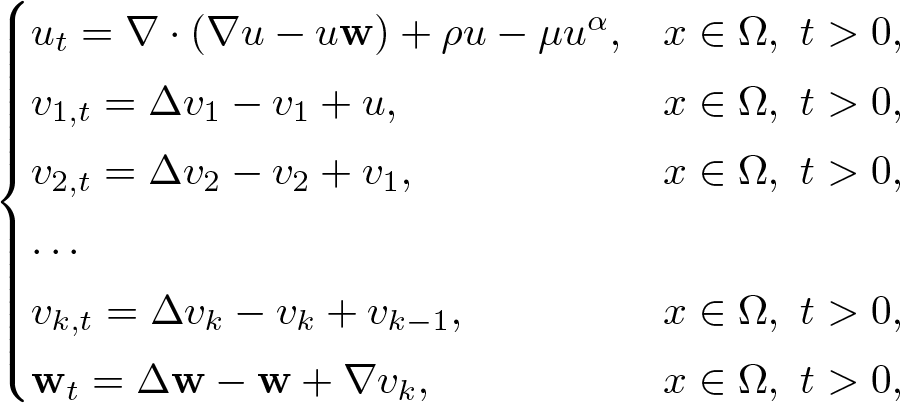 \begin{equation*}\begin{cases}u_t = \nabla\cdot \left(\nabla u - u\bf{w} \right) + \rho u - \mu u^\alpha ,&x \in \Omega ,\ t \gt 0,\\v_{1,t} = \Delta v_1 - v_1 + u,&x \in \Omega ,\ t \gt 0,\\v_{2,t} = \Delta v_2 - v_2 + v_1,&x \in \Omega ,\ t \gt 0,\\\ldots \\v_{k,t} = \Delta v_k - v_k + v_{k - 1},&x \in \Omega ,\ t \gt 0,\\{\bf{w}}_t = \Delta {\bf{w}} - {\bf{w}} + \nabla v_k,&x \in \Omega ,\ t \gt 0,\end{cases}\end{equation*}
\begin{equation*}\begin{cases}u_t = \nabla\cdot \left(\nabla u - u\bf{w} \right) + \rho u - \mu u^\alpha ,&x \in \Omega ,\ t \gt 0,\\v_{1,t} = \Delta v_1 - v_1 + u,&x \in \Omega ,\ t \gt 0,\\v_{2,t} = \Delta v_2 - v_2 + v_1,&x \in \Omega ,\ t \gt 0,\\\ldots \\v_{k,t} = \Delta v_k - v_k + v_{k - 1},&x \in \Omega ,\ t \gt 0,\\{\bf{w}}_t = \Delta {\bf{w}} - {\bf{w}} + \nabla v_k,&x \in \Omega ,\ t \gt 0,\end{cases}\end{equation*}under the homogeneous Neumann boundary condition for u, vi and the homogeneous Dirichlet boundary condition for
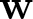 $\bf{w}$ in a smooth bounded domain
$\bf{w}$ in a smooth bounded domain 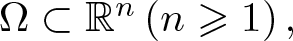 $\Omega \subset {\mathbb{R}^n}\left( {n \geqslant 1} \right),$ where ρ > 0, µ > 0, α > 1 and
$\Omega \subset {\mathbb{R}^n}\left( {n \geqslant 1} \right),$ where ρ > 0, µ > 0, α > 1 and 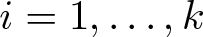 $i=1,\ldots,k$. We reveal that when the index α, the spatial variable n, and the number of equations k satisfy certain relationships, the global solution of the system exists and converges to the constant equilibrium state in the form of exponential convergence.
$i=1,\ldots,k$. We reveal that when the index α, the spatial variable n, and the number of equations k satisfy certain relationships, the global solution of the system exists and converges to the constant equilibrium state in the form of exponential convergence.
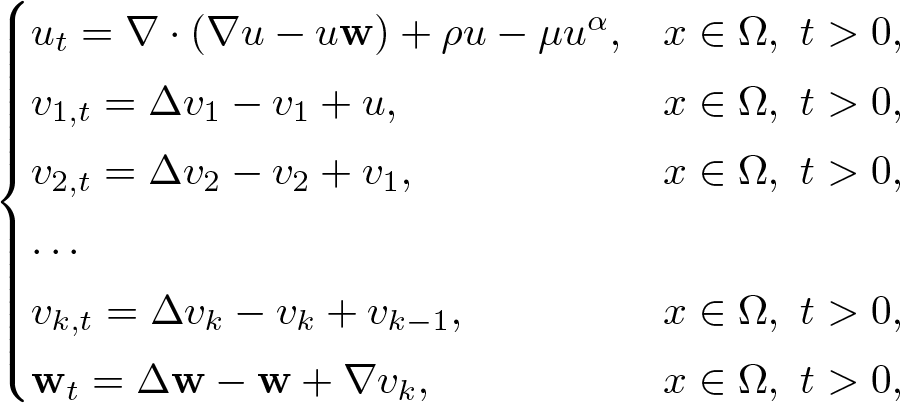
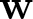
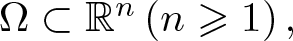
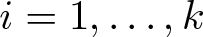
The evolution problem for the 1D nonlocal Fisher-KPP equation with a top hat kernel. Part 2. The initial-boundary value problem on a finite domain
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 04 July 2025, pp. 1-31
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In the second part of this series of papers, we address the same evolution problem that was considered in part 1 (see [16]), namely the nonlocal Fisher-KPP equation in one spatial dimension,
where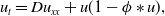 \begin{equation*} u_t = D u_{xx} + u(1-\phi *u), \end{equation*}
\begin{equation*} u_t = D u_{xx} + u(1-\phi *u), \end{equation*}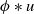 $\phi *u$ is a spatial convolution with the top hat kernel,
$\phi *u$ is a spatial convolution with the top hat kernel, 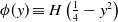 $\phi (y) \equiv H\left (\frac {1}{4}-y^2\right )$, except that now we modify this to an associated initial-boundary value problem on the finite spatial interval
$\phi (y) \equiv H\left (\frac {1}{4}-y^2\right )$, except that now we modify this to an associated initial-boundary value problem on the finite spatial interval 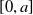 $[0,a]$ rather than the whole real line. Boundary conditions are required at the end points of the interval, and we address the situations when these are of either Dirichlet or Neumann type. This model is a natural extension of the classical Fisher-KPP model, with the introduction of the simplest possible nonlocal effect into the saturation term. Nonlocal reaction-diffusion models arise naturally in a variety of (frequently biological or ecological) contexts, and as such it is of fundamental interest to examine their properties in detail, and to compare and contrast these with the well known properties of the classical Fisher-KPP model.
$[0,a]$ rather than the whole real line. Boundary conditions are required at the end points of the interval, and we address the situations when these are of either Dirichlet or Neumann type. This model is a natural extension of the classical Fisher-KPP model, with the introduction of the simplest possible nonlocal effect into the saturation term. Nonlocal reaction-diffusion models arise naturally in a variety of (frequently biological or ecological) contexts, and as such it is of fundamental interest to examine their properties in detail, and to compare and contrast these with the well known properties of the classical Fisher-KPP model.
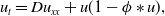
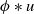
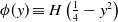
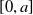
Cahn–Hilliard equations with singular potential, reaction term and pure phase initial datum
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 27 May 2025, pp. 1-26
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Collapsing immortal Kähler-Ricci flows
- Part of
-
- Journal:
- Forum of Mathematics, Pi / Volume 13 / 2025
- Published online by Cambridge University Press:
- 19 May 2025, e18
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
DELAYED HOPF BIFURCATIONS IN REACTION–DIFFUSION SYSTEMS IN TWO SPACE DIMENSIONS
- Part of
-
- Journal:
- The ANZIAM Journal / Volume 67 / 2025
- Published online by Cambridge University Press:
- 13 May 2025, e19
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Global dynamics for the generalised chemotaxis-Navier–Stokes system in
 $\mathbb{R}^3$
$\mathbb{R}^3$
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 22 April 2025, pp. 1-38
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider the chemotaxis-Navier–Stokes system with generalised fluid dissipation in
 $\mathbb{R}^3$:which models the motion of swimming bacteria in water flows. First, we prove blow-up criteria of strong solutions to the Cauchy problem, including the Prodi-Serrin-type criterion for
$\mathbb{R}^3$:which models the motion of swimming bacteria in water flows. First, we prove blow-up criteria of strong solutions to the Cauchy problem, including the Prodi-Serrin-type criterion for \begin{eqnarray*} \begin{cases} \partial _t n+u\cdot \nabla n=\Delta n- \nabla \cdot (\chi (c)n \nabla c),\\[5pt] \partial _t c+u \cdot \nabla c=\Delta c-nf(c),\\[5pt] \partial _t u +u \cdot \nabla u+\nabla P=-(\!-\Delta )^\alpha u-n\nabla \phi, \\[5pt] \nabla \cdot u=0, \end{cases} \end{eqnarray*}
\begin{eqnarray*} \begin{cases} \partial _t n+u\cdot \nabla n=\Delta n- \nabla \cdot (\chi (c)n \nabla c),\\[5pt] \partial _t c+u \cdot \nabla c=\Delta c-nf(c),\\[5pt] \partial _t u +u \cdot \nabla u+\nabla P=-(\!-\Delta )^\alpha u-n\nabla \phi, \\[5pt] \nabla \cdot u=0, \end{cases} \end{eqnarray*} $\alpha \gt \frac {3}{4}$ and the Beir
$\alpha \gt \frac {3}{4}$ and the Beir $\tilde {\textrm {a}}$o da Veiga-type criterion for
$\tilde {\textrm {a}}$o da Veiga-type criterion for  $\alpha \gt \frac {1}{2}$. Then, we verify the global existence and uniqueness of strong solutions for arbitrarily large initial fluid velocity and bacteria density for
$\alpha \gt \frac {1}{2}$. Then, we verify the global existence and uniqueness of strong solutions for arbitrarily large initial fluid velocity and bacteria density for  $\alpha \geq \frac {5}{4}$. Furthermore, in the scenario of
$\alpha \geq \frac {5}{4}$. Furthermore, in the scenario of  $\frac {3}{4}\lt \alpha \lt \frac {5}{4}$, we establish uniform regularity estimates and optimal time-decay rates of global solutions if only the
$\frac {3}{4}\lt \alpha \lt \frac {5}{4}$, we establish uniform regularity estimates and optimal time-decay rates of global solutions if only the  $L^2$-norm of initial data is small. To our knowledge, this work provides the first result concerning the global existence and large-time behaviour of strong solutions for the chemotaxis-Navier–Stokes equations with possibly large oscillations.
$L^2$-norm of initial data is small. To our knowledge, this work provides the first result concerning the global existence and large-time behaviour of strong solutions for the chemotaxis-Navier–Stokes equations with possibly large oscillations.









Pattern formation with jump discontinuity in a predator–prey model with Holling-II functional response
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 11 April 2025, pp. 1160-1182
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Spatio-temporal dynamics of a lattice prey–predator system with non-local diffusion in a periodic habitat
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 31 March 2025, pp. 1-43
-
- Article
- Export citation
Parabolic Muckenhoupt weights characterized by parabolic fractional maximal and integral operators with time lag
- Part of
-
- Journal:
- Canadian Journal of Mathematics , First View
- Published online by Cambridge University Press:
- 17 March 2025, pp. 1-54
-
- Article
- Export citation
Impact of prey-taxis on a harvested intraguild predation predator–prey model
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 10 March 2025, pp. 1183-1220
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
A critical non-homogeneous heat equation with weighted source
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 06 March 2025, pp. 1148-1159
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Some qualitative properties of radially symmetric solutions to the non-homogeneous heat equation with critical density and weighted source
are obtained, in the range of exponents \begin{align*} |x|^{-2}\partial _tu=\Delta u+|x|^{\sigma }u^p, \quad (x,t)\in {\mathbb {R}}^N\times (0,T), \end{align*}
\begin{align*} |x|^{-2}\partial _tu=\Delta u+|x|^{\sigma }u^p, \quad (x,t)\in {\mathbb {R}}^N\times (0,T), \end{align*} $p\gt 1$,
$p\gt 1$,  $\sigma \ge -2$. More precisely, we establish conditions fulfilled by the initial data in order for the solutions to either blow-up in finite time or decay to zero as
$\sigma \ge -2$. More precisely, we establish conditions fulfilled by the initial data in order for the solutions to either blow-up in finite time or decay to zero as  $t\to \infty$ and, in the latter case, we also deduce decay rates and large time behavior. In the limiting case
$t\to \infty$ and, in the latter case, we also deduce decay rates and large time behavior. In the limiting case  $\sigma =-2$, we prove the existence of non-trivial, non-negative solutions, in stark contrast to the homogeneous case. A transformation to a generalized Fisher–KPP equation is derived and employed in order to deduce these properties.
$\sigma =-2$, we prove the existence of non-trivial, non-negative solutions, in stark contrast to the homogeneous case. A transformation to a generalized Fisher–KPP equation is derived and employed in order to deduce these properties.





Global well-posedness and Turing–Hopf bifurcation of prey-taxis systems with hunting cooperation
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 6 / December 2025
- Published online by Cambridge University Press:
- 24 February 2025, pp. 1121-1147
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Singular SPDEs on homogeneous lie groups
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 18 February 2025, pp. 1-72
-
- Article
- Export citation
-
The aim of this article is to extend the scope of the theory of regularity structures in order to deal with a large class of singular stochastic partial differential equations of the form
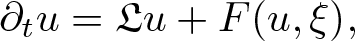 \begin{equation*}\partial_t u = \mathfrak{L} u+ F(u, \xi),\end{equation*}
\begin{equation*}\partial_t u = \mathfrak{L} u+ F(u, \xi),\end{equation*}where the differential operator
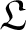 $\mathfrak{L}$ fails to be elliptic. This is achieved by interpreting the base space
$\mathfrak{L}$ fails to be elliptic. This is achieved by interpreting the base space 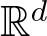 $\mathbb{R}^{d}$ as a non-trivial homogeneous Lie group
$\mathbb{R}^{d}$ as a non-trivial homogeneous Lie group 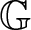 $\mathbb{G}$ such that the differential operator
$\mathbb{G}$ such that the differential operator 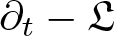 $\partial_t -\mathfrak{L}$ becomes a translation invariant hypoelliptic operator on
$\partial_t -\mathfrak{L}$ becomes a translation invariant hypoelliptic operator on 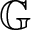 $\mathbb{G}$. Prime examples are the kinetic Fokker-Planck operator
$\mathbb{G}$. Prime examples are the kinetic Fokker-Planck operator 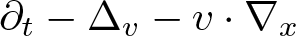 $\partial_t -\Delta_v - v\cdot \nabla_x$ and heat-type operators associated with sub-Laplacians. As an application of the developed framework, we solve a class of parabolic Anderson type equations
$\partial_t -\Delta_v - v\cdot \nabla_x$ and heat-type operators associated with sub-Laplacians. As an application of the developed framework, we solve a class of parabolic Anderson type equations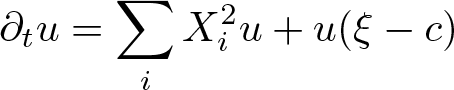 \begin{equation*}\partial_t u = \sum_{i} X^2_i u + u (\xi-c)\end{equation*}
\begin{equation*}\partial_t u = \sum_{i} X^2_i u + u (\xi-c)\end{equation*}on the compact quotient of an arbitrary Carnot group.
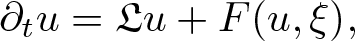
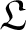
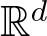
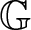
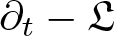
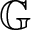
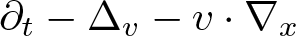
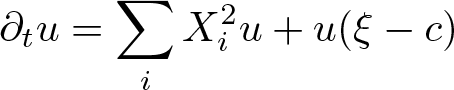
Continuous in time bubble decomposition for the harmonic map heat flow
- Part of
-
- Journal:
- Forum of Mathematics, Pi / Volume 13 / 2025
- Published online by Cambridge University Press:
- 11 February 2025, e4
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider the harmonic map heat flow for maps
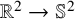 $\mathbb {R}^{2} \to \mathbb {S}^2$. It is known that solutions to the initial value problem exhibit bubbling along a well-chosen sequence of times. We prove that every sequence of times admits a subsequence along which bubbling occurs. This is deduced as a corollary of our main theorem, which shows that the solution approaches the family of multi-bubble configurations in continuous time.
$\mathbb {R}^{2} \to \mathbb {S}^2$. It is known that solutions to the initial value problem exhibit bubbling along a well-chosen sequence of times. We prove that every sequence of times admits a subsequence along which bubbling occurs. This is deduced as a corollary of our main theorem, which shows that the solution approaches the family of multi-bubble configurations in continuous time.
Lyapunov stability of non-isolated equilibria for strongly irreversible Allen–Cahn equations
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 18 December 2024, pp. 1-21
-
- Article
- Export citation
Fractional time differential equations as a singular limit of the Kobayashi–Warren–Carter system
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 11 December 2024, pp. 1-37
-
- Article
- Export citation
Analysis of a model describing bacterial colony expansion in radial geometry driven by chemotaxis
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 5 / October 2025
- Published online by Cambridge University Press:
- 02 December 2024, pp. 910-947
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Well-posedness of quasilinear parabolic equations in time-weighted spaces
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 26 November 2024, pp. 1-33
-
- Article
- Export citation
-
Well-posedness in time-weighted spaces of certain quasilinear (and semilinear) parabolic evolution equations
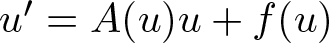 $u'=A(u)u+f(u)$ is established. The focus lies on the case of strict inclusions
$u'=A(u)u+f(u)$ is established. The focus lies on the case of strict inclusions 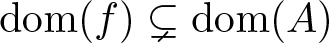 $\mathrm{dom}(f)\subsetneq \mathrm{dom}(A)$ of the domains of the nonlinearities
$\mathrm{dom}(f)\subsetneq \mathrm{dom}(A)$ of the domains of the nonlinearities 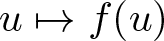 $u\mapsto f(u)$ and
$u\mapsto f(u)$ and 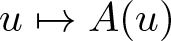 $u\mapsto A(u)$. Based on regularizing effects of parabolic equations it is shown that a semiflow is generated in intermediate spaces. In applications this allows one to derive global existence from weaker a priori estimates. The result is illustrated by examples of chemotaxis systems.
$u\mapsto A(u)$. Based on regularizing effects of parabolic equations it is shown that a semiflow is generated in intermediate spaces. In applications this allows one to derive global existence from weaker a priori estimates. The result is illustrated by examples of chemotaxis systems.
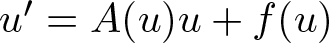
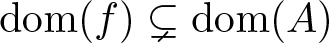
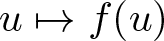
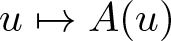
Asymptotic estimate of solutions in a 4th-order parabolic equation with the Frobenius norm of a Hessian matrix
- Part of
-
- Journal:
- Canadian Mathematical Bulletin / Volume 68 / Issue 1 / March 2025
- Published online by Cambridge University Press:
- 25 November 2024, pp. 91-108
- Print publication:
- March 2025
-
- Article
- Export citation
Global well-posedness and uniform boundedness of 2D urban crime models with nonlinear advection enhancement
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 5 / October 2025
- Published online by Cambridge University Press:
- 20 November 2024, pp. 897-909
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
