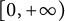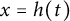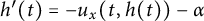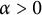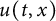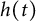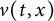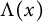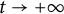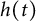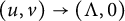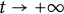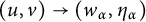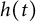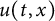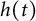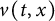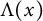Refine listing
Actions for selected content:
308 results in 35Kxx
Threshold dynamics of a time-periodic SIS model with anomalous diffusion in heterogeneous environments
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 24 February 2026, pp. 1-28
-
- Article
- Export citation
Proliferation-dominated large time behaviour in a planar doubly degenerate nutrient taxis system
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 23 February 2026, pp. 1-30
-
- Article
- Export citation
-
This study is concerned with nonnegative solutions of the no-flux initial-boundary value problem for the doubly degenerate nutrient taxis system with a logistic source, as given by
in a smoothly bounded planar domain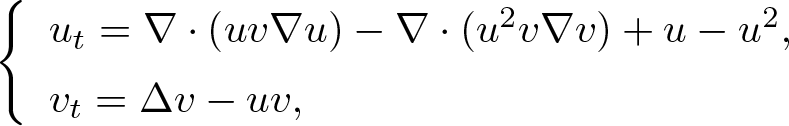 \begin{align*}\left\{\begin{array}{ll}u_t = \nabla\cdot (uv\nabla u) - \nabla\cdot (u^2 v\nabla v)+ u - u^2,\\[1mm]v_t = \Delta v -uv,\end{array} \right.\end{align*}
\begin{align*}\left\{\begin{array}{ll}u_t = \nabla\cdot (uv\nabla u) - \nabla\cdot (u^2 v\nabla v)+ u - u^2,\\[1mm]v_t = \Delta v -uv,\end{array} \right.\end{align*} $\Omega$ with
$\Omega$ with 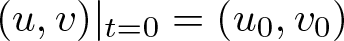 $(u,v)|_{t=0}=(u_0,v_0)$. It is shown that despite substantially weakened diffusion, the stabilizing effects of logistic-type cell kinetics may overbalance any heterogeneity-supporting tendency of cross-diffusion: Namely, it is seen that for all suitably smooth initial data satisfying
$(u,v)|_{t=0}=(u_0,v_0)$. It is shown that despite substantially weakened diffusion, the stabilizing effects of logistic-type cell kinetics may overbalance any heterogeneity-supporting tendency of cross-diffusion: Namely, it is seen that for all suitably smooth initial data satisfying 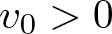 $v_0 \gt 0$ in
$v_0 \gt 0$ in  $\overline{\Omega}$ anda corresponding continuous weak solution, known to exist globally according to previous literature, has the property that as
$\overline{\Omega}$ anda corresponding continuous weak solution, known to exist globally according to previous literature, has the property that as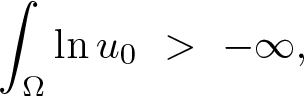 \begin{align*}\int_\Omega \ln u_0~ \gt ~-\infty,\end{align*}
\begin{align*}\int_\Omega \ln u_0~ \gt ~-\infty,\end{align*}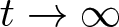 $t\to\infty$,
$t\to\infty$,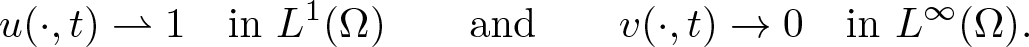 \begin{align*} u(\cdot,t) \rightharpoonup 1 \quad \text{in } L^1(\Omega) \qquad\text{and}\qquad v(\cdot,t)\to 0 \quad \text{in } L^\infty(\Omega). \end{align*}
\begin{align*} u(\cdot,t) \rightharpoonup 1 \quad \text{in } L^1(\Omega) \qquad\text{and}\qquad v(\cdot,t)\to 0 \quad \text{in } L^\infty(\Omega). \end{align*}
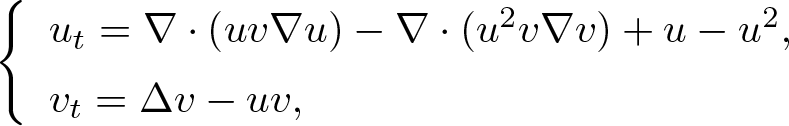

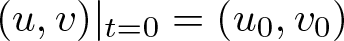
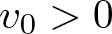

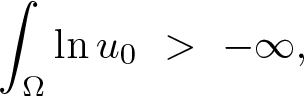
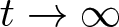
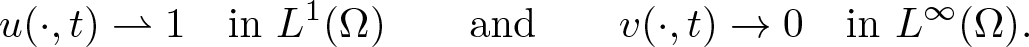
Periodic dynamics in a time-switching advection-diffusion model for competing Aedes mosquitoes
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 16 February 2026, pp. 1-21
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
RENORMALIZED AND ENTROPY SOLUTIONS TO THE GENERAL NONLINEAR PARABOLIC EQUATIONS IN MUSIELAK–ORLICZ SPACES
- Part of
-
- Journal:
- Journal of the Australian Mathematical Society , First View
- Published online by Cambridge University Press:
- 30 January 2026, pp. 1-30
-
- Article
- Export citation
Competition of small targets in planar domains: from Dirichlet to Robin and Steklov boundary condition
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 12 January 2026, pp. 1-48
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Rigidity for the heat equation with density on Riemannian manifolds through a conformal change
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 12 January 2026, pp. 1-41
-
- Article
- Export citation
-
We investigate uniqueness of solution to the heat equation with a density
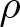 $\rho$ on complete, non-compact weighted Riemannian manifolds of infinite volume. Our main goal is to identify sufficient conditions under which the solution
$\rho$ on complete, non-compact weighted Riemannian manifolds of infinite volume. Our main goal is to identify sufficient conditions under which the solution 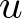 $u$ vanishes identically, assuming that
$u$ vanishes identically, assuming that 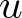 $u$ belongs to a certain weighted Lebesgue space with exponential or polynomial weight,
$u$ belongs to a certain weighted Lebesgue space with exponential or polynomial weight, 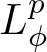 $L^p_{\phi}$. We distinguish between the cases
$L^p_{\phi}$. We distinguish between the cases 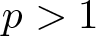 $p \gt 1$ and
$p \gt 1$ and 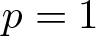 $p = 1$ which required stronger assumptions on the manifold and the density function
$p = 1$ which required stronger assumptions on the manifold and the density function 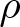 $\rho$. We develop a unified method based on a conformal transformation of the metric, which allows us to reduce the problem to a standard heat equation on a suitably weighted manifold. In addition, we construct explicit counterexamples on model manifolds which demonstrate optimality of our assumptions on the density
$\rho$. We develop a unified method based on a conformal transformation of the metric, which allows us to reduce the problem to a standard heat equation on a suitably weighted manifold. In addition, we construct explicit counterexamples on model manifolds which demonstrate optimality of our assumptions on the density 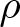 $\rho$.
$\rho$.
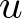
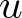
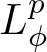
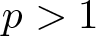
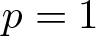
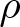
A note on time-asymptotic bounds with a sharp algebraic rate and a transitional exponent for the sublinear Fujita problem
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 08 January 2026, pp. 1-10
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Propagation dynamics of time-dependent reaction-diffusion equations under climate change in an infinite cylinder
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 19 December 2025, pp. 1-22
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In this paper, we consider a reaction-diffusion equation that models the time-almost periodic response to climate change within a straight, infinite cylindrical domain. The shifting edge of the habitat is characterised by a time-almost periodic function, reflecting the varying pace of environmental changes. Note that the principal spectral theory is an important role to study the dynamics of reaction-diffusion equations in time heterogeneous environment. Initially, for time-almost periodic parabolic equations in finite cylindrical domains, we develop the principal spectral theory of such equations with mixed Dirichlet–Neumann boundary conditions. Subsequently, we demonstrate that the approximate principal Lyapunov exponent serves as a definitive threshold for species persistence versus extinction. Then, the existence, exponential decay and stability of the forced wave solutions
 $U(t,x_{1},y)=V\left (t,x_{1}-\int ^{t}_{0}c(s)ds,y\right )$ are established. Additionally, we analyse how fluctuations in the shifting speed affect the approximate top Lyapunov exponent.
$U(t,x_{1},y)=V\left (t,x_{1}-\int ^{t}_{0}c(s)ds,y\right )$ are established. Additionally, we analyse how fluctuations in the shifting speed affect the approximate top Lyapunov exponent.

On a Cahn–Hilliard equation for the growth and division of chemically active droplets modelling protocells
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 06 November 2025, pp. 1-31
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Impact of birth pulse and environment shift on population survival and propagation
- Part of
-
- Journal:
- Canadian Journal of Mathematics , First View
- Published online by Cambridge University Press:
- 17 October 2025, pp. 1-27
-
- Article
- Export citation
A remark on the long-time asymptotics of global-in-time solutions for semilinear parabolic equations in
 ${\mathbb {R}^N}$
${\mathbb {R}^N}$
- Part of
-
- Journal:
- Canadian Journal of Mathematics , First View
- Published online by Cambridge University Press:
- 10 October 2025, pp. 1-21
-
- Article
- Export citation
-
We consider the asymptotics of long-time behavior of a solution u of the semilinear parabolic problem
 $\partial _tu=\Delta u-u+u|u|^{p-2}$ in
$\partial _tu=\Delta u-u+u|u|^{p-2}$ in  ${\mathbb {R}^N}\times (0,\infty )$,
${\mathbb {R}^N}\times (0,\infty )$,  $u(0)=u_0\in H^1({\mathbb {R}^N})\cap L^\infty ({\mathbb {R}^N})$. Since the spatial domain on which the problem is posed is noncompact, we cannot expect the relative compactness of the solution orbit, e.g., in
$u(0)=u_0\in H^1({\mathbb {R}^N})\cap L^\infty ({\mathbb {R}^N})$. Since the spatial domain on which the problem is posed is noncompact, we cannot expect the relative compactness of the solution orbit, e.g., in  $H^1({\mathbb {R}^N})$ in general. In this article, we prove that the compactness of the orbit holds up to the ground state energy level, namely, if
$H^1({\mathbb {R}^N})$ in general. In this article, we prove that the compactness of the orbit holds up to the ground state energy level, namely, if  $\lim _{t\to \infty }I(u(t))\leq d_\infty $, where I is the energy functional associated with (P) and
$\lim _{t\to \infty }I(u(t))\leq d_\infty $, where I is the energy functional associated with (P) and  $d_\infty $ its ground state energy, then the orbit of
$d_\infty $ its ground state energy, then the orbit of  $u(t)$ is compact in
$u(t)$ is compact in  $H^1({\mathbb {R}^N})$. Our result includes the previous results in [4, 5].
$H^1({\mathbb {R}^N})$. Our result includes the previous results in [4, 5].









Nonlinear Hodge flows in symplectic geometry
- Part of
-
- Journal:
- Compositio Mathematica / Volume 161 / Issue 10 / October 2025
- Published online by Cambridge University Press:
- 17 December 2025, pp. 2461-2492
- Print publication:
- October 2025
-
- Article
- Export citation
-
Given a symplectic class
 $$\left[ \omega \right]$$ on a four torus
$$\left[ \omega \right]$$ on a four torus 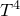 $${T^4}$$ (or a
$${T^4}$$ (or a 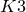 $$K3$$ surface), a folklore problem in symplectic geometry is whether symplectic forms in
$$K3$$ surface), a folklore problem in symplectic geometry is whether symplectic forms in  $$\left[ \omega \right]$$ are isotopic to each other. We introduce a family of nonlinear Hodge heat flows on compact symplectic four manifolds to approach this problem, which is an adaption of nonlinear Hodge theory in symplectic geometry. As a particular example, we study a conformal Hodge heat flow in detail. We prove a stability result of the flow near an almost Kähler structure
$$\left[ \omega \right]$$ are isotopic to each other. We introduce a family of nonlinear Hodge heat flows on compact symplectic four manifolds to approach this problem, which is an adaption of nonlinear Hodge theory in symplectic geometry. As a particular example, we study a conformal Hodge heat flow in detail. We prove a stability result of the flow near an almost Kähler structure 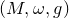 $$\left( {M,\omega ,g} \right)$$. We also prove that, if
$$\left( {M,\omega ,g} \right)$$. We also prove that, if 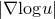 $$\left| {\nabla {\rm{log}}u} \right|$$ stays bounded along the flow, then the flow exists for all time for any initial symplectic form
$$\left| {\nabla {\rm{log}}u} \right|$$ stays bounded along the flow, then the flow exists for all time for any initial symplectic form 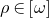 $$\rho \in \left[ \omega \right]$$ and it converges to
$$\rho \in \left[ \omega \right]$$ and it converges to  $$\omega $$ smoothly along the flow with uniform control, where
$$\omega $$ smoothly along the flow with uniform control, where 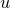 $$u$$ is the volume potential of
$$u$$ is the volume potential of 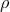 $$\rho $$.
$$\rho $$.


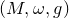
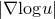
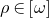

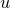
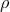
Shrinking vs. expanding: the evolution of spatial support in degenerate Keller–Segel systems
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 30 September 2025, pp. 1-25
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We consider radially symmetric solutions of the degenerate Keller–Segel system
 \begin{align*}\begin{cases}\partial_t u=\nabla\cdot (u^{m-1}\nabla u - u\nabla v),\\0=\Delta v -\mu +u,\quad\mu =\frac{1}{|\Omega|}\int_\Omega u,\end{cases}\end{align*}
\begin{align*}\begin{cases}\partial_t u=\nabla\cdot (u^{m-1}\nabla u - u\nabla v),\\0=\Delta v -\mu +u,\quad\mu =\frac{1}{|\Omega|}\int_\Omega u,\end{cases}\end{align*}in balls
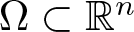 $\Omega\subset\mathbb R^n$,
$\Omega\subset\mathbb R^n$, 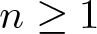 $n\ge 1$, where m > 1 is arbitrary. Our main result states that the initial evolution of the positivity set of u is essentially determined by the shape of the (nonnegative, radially symmetric, Hölder continuous) initial data u0 near the boundary of its support
$n\ge 1$, where m > 1 is arbitrary. Our main result states that the initial evolution of the positivity set of u is essentially determined by the shape of the (nonnegative, radially symmetric, Hölder continuous) initial data u0 near the boundary of its support 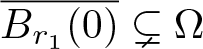 $\overline{B_{r_1}(0)}\subsetneq\Omega$: It shrinks for sufficiently flat and expands for sufficiently steep u0. More precisely, there exists an explicit constant
$\overline{B_{r_1}(0)}\subsetneq\Omega$: It shrinks for sufficiently flat and expands for sufficiently steep u0. More precisely, there exists an explicit constant 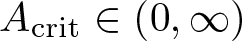 $A_{\mathrm{crit}} \in (0, \infty)$ (depending only on
$A_{\mathrm{crit}} \in (0, \infty)$ (depending only on 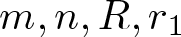 $m, n, R, r_1$ and
$m, n, R, r_1$ and  $\int_\Omega u_0$) such that if
$\int_\Omega u_0$) such that if 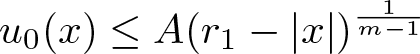 $u_0(x)\le A(r_1-|x|)^\frac{1}{m-1}$ for all
$u_0(x)\le A(r_1-|x|)^\frac{1}{m-1}$ for all  $|x|\in(r_0, r_1)$ and some
$|x|\in(r_0, r_1)$ and some  $r_0\in(0,r_1)$ and
$r_0\in(0,r_1)$ and 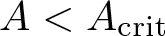 $A \lt A_{\mathrm{crit}}$ then there are T > 0 and ζ > 0 such that
$A \lt A_{\mathrm{crit}}$ then there are T > 0 and ζ > 0 such that 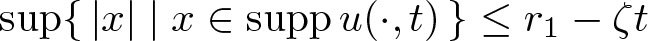 $\sup\{\, |x| \mid x \in \operatorname{supp} u(\cdot, t)\,\}\le r_1 -\zeta t$ for all
$\sup\{\, |x| \mid x \in \operatorname{supp} u(\cdot, t)\,\}\le r_1 -\zeta t$ for all 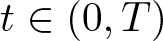 $t\in(0, T)$, while if
$t\in(0, T)$, while if 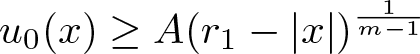 $u_0(x)\ge A(r_1-|x|)^\frac{1}{m-1}$ for all
$u_0(x)\ge A(r_1-|x|)^\frac{1}{m-1}$ for all 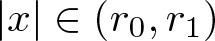 $|x|\in(r_0, r_1)$ and some
$|x|\in(r_0, r_1)$ and some 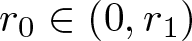 $r_0 \in (0, r_1)$ and
$r_0 \in (0, r_1)$ and  $A \gt A_{\mathrm{crit}}$ then we can find T > 0 and ζ > 0 such that
$A \gt A_{\mathrm{crit}}$ then we can find T > 0 and ζ > 0 such that 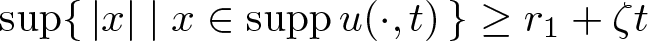 $\sup\{\, |x| \mid x \in \operatorname{supp} u(\cdot, t)\,\}\ge r_1 +\zeta t$ for all
$\sup\{\, |x| \mid x \in \operatorname{supp} u(\cdot, t)\,\}\ge r_1 +\zeta t$ for all 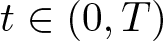 $t\in(0, T)$.
$t\in(0, T)$.

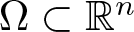
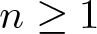
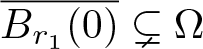
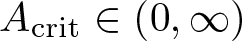
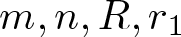

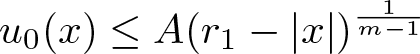


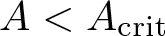
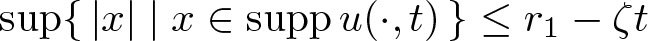
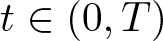
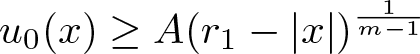
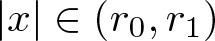
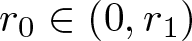

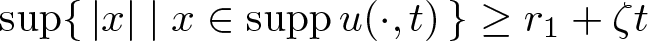
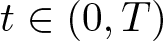
Anisotropic area-preserving flows for plane curves and entropy inequalities
- Part of
-
- Journal:
- Canadian Journal of Mathematics , First View
- Published online by Cambridge University Press:
- 23 September 2025, pp. 1-25
-
- Article
- Export citation
Global dynamics of a degenerate reaction-diffusion cholera model with phage-bacteria interaction in a heterogeneous environment
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 22 September 2025, pp. 1-32
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
To investigate multiple effects of the interaction between V. cholerae and phage on cholera transmission, we propose a degenerate reaction-diffusion model with different dispersal rates, which incorporates a short-lived hyperinfectious (HI vibrios) state of V. cholerae and lower-infectious (LI vibrios) state of V. cholerae. Our main purpose is to investigate the existence and stability analysis of multi-class boundary steady states, which is much more complicated and challenging than the case when the boundary steady state is unique. In a spatially heterogeneous case, the basic reproduction number
 $\mathscr{R}_{0}$ is defined as the spectral radius of the sum of two linear operators associated with HI vibrios infection and LI vibrios infection. If
$\mathscr{R}_{0}$ is defined as the spectral radius of the sum of two linear operators associated with HI vibrios infection and LI vibrios infection. If  $\mathscr{R}_{0}\leq 1$, the disease-free steady state is globally asymptotically stable. If
$\mathscr{R}_{0}\leq 1$, the disease-free steady state is globally asymptotically stable. If  $\mathscr{R}_{0}\gt 1$, the uniform persistence of phage-free model, as well as the existence of the phage-free steady state, are established. In a spatially homogeneous case, when
$\mathscr{R}_{0}\gt 1$, the uniform persistence of phage-free model, as well as the existence of the phage-free steady state, are established. In a spatially homogeneous case, when  $\ \;\widetilde{\!\!\!\mathscr{R}}_{0}\gt 1$, the global asymptotic stability of phage-free steady state and the uniform persistence of the phage-present model are discussed under some additional conditions. The mathematical approach here has wide applications in degenerate Partial Differential Equations.
$\ \;\widetilde{\!\!\!\mathscr{R}}_{0}\gt 1$, the global asymptotic stability of phage-free steady state and the uniform persistence of the phage-present model are discussed under some additional conditions. The mathematical approach here has wide applications in degenerate Partial Differential Equations.




On the convergence of non-integer linear Hopf flow
- Part of
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 69 / Issue 1 / February 2026
- Published online by Cambridge University Press:
- 12 September 2025, pp. 255-283
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Global existence and some qualitative properties of weak solutions for a class of pseudo-parabolic equations with a logarithmic nonlinearity in whole
 $\mathbb{R}^{N}$
$\mathbb{R}^{N}$
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 01 August 2025, pp. 1-44
-
- Article
- Export citation
-
In this paper, we study the Cauchy problem for pseudo-parabolic equations with a logarithmic nonlinearity. After establishing the existence and uniqueness of weak solutions within a suitable functional framework, we investigate several qualitative properties, including the asymptotic behaviour and blow-up of solutions as
 $t\to +\infty$. Moreover, when the initial data are close to a Gaussian function, we prove that these weak solutions exhibit either super-exponential growth or super-exponential decay.
$t\to +\infty$. Moreover, when the initial data are close to a Gaussian function, we prove that these weak solutions exhibit either super-exponential growth or super-exponential decay.


On a Santaló point for Nakamura-Tsuji’s Laplace transform inequality
- Part of
-
- Journal:
- Forum of Mathematics, Sigma / Volume 13 / 2025
- Published online by Cambridge University Press:
- 24 July 2025, e125
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Existence-uniqueness theory and small-data decay for a reaction-diffusion model of wildfire spread
- Part of
-
- Journal:
- Canadian Mathematical Bulletin / Volume 68 / Issue 4 / December 2025
- Published online by Cambridge University Press:
- 08 July 2025, pp. 1347-1358
- Print publication:
- December 2025
-
- Article
- Export citation
-
I examine some analytical properties of a nonlinear reaction-diffusion system that has been used to model the propagation of a wildfire. I establish global-in-time existence and uniqueness of bounded mild solutions to the Cauchy problem for this system given bounded initial data. In particular, this shows that the model does not allow for thermal blow-up. If the initial temperature and fuel density also satisfy certain integrability conditions, the spatial
 $L^2$-norms of these global solutions are uniformly bounded in time. Additionally, I use a bootstrap argument to show that small initial temperatures give rise to solutions that decay to zero as time goes to infinity, proving the existence of initial states that do not develop into traveling combustion waves.
$L^2$-norms of these global solutions are uniformly bounded in time. Additionally, I use a bootstrap argument to show that small initial temperatures give rise to solutions that decay to zero as time goes to infinity, proving the existence of initial states that do not develop into traveling combustion waves.

A free boundary problem of competition-diffusion system with Dirichlet boundary condition
- Part of
-
- Journal:
- Canadian Mathematical Bulletin / Volume 69 / Issue 1 / March 2026
- Published online by Cambridge University Press:
- 07 July 2025, pp. 1-20
- Print publication:
- March 2026
-
- Article
- Export citation
-
In this article, we investigate a free boundary problem for the Lotka–Volterra model consisting of an invasive species with density u and a native species with density v in one dimension. We assume that v undergoes diffusion and growth in
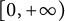 $[0,+\infty )$, and u invades into the environment with spreading front
$[0,+\infty )$, and u invades into the environment with spreading front 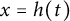 $x=h(t)$ satisfying free boundary condition
$x=h(t)$ satisfying free boundary condition 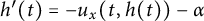 $h'(t)=-u_x(t,h(t))-\alpha $ for some decay rate
$h'(t)=-u_x(t,h(t))-\alpha $ for some decay rate 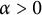 $\alpha>0$, this is caused by the bad environment at the boundary. When u is an inferior competitor,
$\alpha>0$, this is caused by the bad environment at the boundary. When u is an inferior competitor, 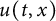 $u(t,x)$ and
$u(t,x)$ and 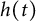 $h(t)$ tend to 0 within a finite time, while another specie
$h(t)$ tend to 0 within a finite time, while another specie 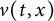 $v(t,x)$ tends to a stationary
$v(t,x)$ tends to a stationary 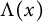 $\Lambda (x)$ defined on the half-line. When u is a superior competitor, we have a trichotomy result: spreading of u and vanishing of v (i.e., as
$\Lambda (x)$ defined on the half-line. When u is a superior competitor, we have a trichotomy result: spreading of u and vanishing of v (i.e., as 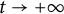 $t \to +\infty $,
$t \to +\infty $, 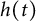 $h(t)$ goes to
$h(t)$ goes to 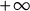 $+\infty $ and
$+\infty $ and 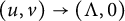 $(u,v)\to (\Lambda ,0)$); the transition case (i.e., as
$(u,v)\to (\Lambda ,0)$); the transition case (i.e., as 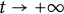 $t \to +\infty $,
$t \to +\infty $, 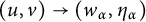 $(u,v)\to (w_\alpha , \eta _\alpha )$,
$(u,v)\to (w_\alpha , \eta _\alpha )$, 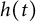 $h(t)$ tends to a finite point); vanishing of u and spreading of v (i.e.,
$h(t)$ tends to a finite point); vanishing of u and spreading of v (i.e., 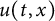 $u(t,x)$ and
$u(t,x)$ and 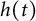 $h(t)$ tends to 0 within a finite time,
$h(t)$ tends to 0 within a finite time, 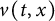 $v(t,x)$ converges to
$v(t,x)$ converges to 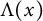 $\Lambda (x)$). Additionally, we show that this trichotomy result depends on the initial data
$\Lambda (x)$). Additionally, we show that this trichotomy result depends on the initial data  $u(0,x)$.
$u(0,x)$.